
JOURNAL OF NATURAL RESOURCES >
Effects of groundwater salinities on photosynthesis and water consumption characteristics of Tamarix chinensis in the Yellow River Delta
Received date: 2019-02-26
Request revised date: 2019-06-22
Online published: 2019-12-28
Copyright
Tamarix chinensis Lour. is a constructive species of the saline-alkali wetland in the Yellow River Delta, and shallow groundwater is the main water source for the growth of T. chinensis in this area. To explore the response mechanism of T. chinensis to groundwater salinity in shallow groundwater area in the Yellow River Delta, parameters of photosynthesis and water consumption were determined under 4 groundwater salinity gradients of fresh (0 g·L-1), brackish (3 g·L-1), saline (8 g·L-1) and salt (20 g·L-1) water at simulated groundwater level of 0.9 m via leaf gas-exchange and stem sap flow (SF) techniques. The results showed that: (1) With increasing groundwater salinity, soil water content, soil salt content and absolute concentration of soil solution increased gradually; (2) Parameters of the max net photosynthetic rate (Pnmax), apparent quantum yield (AQY), light saturation point (LSP), stomatal conductivity (Gs), transpiration rate (Tr), intercellular CO2 concentration (Ci) and stem SF rate increased first and then decreased with the increase of groundwater salinities, reaching the maximum values respectively under brackish groundwater (Gs, AQY, Tr, Ci, and SF rate) and saline groundwater (Pnmax and LSP) treatments, while all values of the above parameters reached the minimum values under salt groundwater treatment; (3) With the increase of groundwater salinity, the water use efficiency (WUE) and stomatal limitation value (Ls) of T. chinensis decreased before increasing, achieving the lowest level under brackish groundwater and the highest under salt groundwater. Under the condition of groundwater depth of 0.9 m, the salinity of groundwater significantly affects the characteristics of soil water and salt contents. The increase of soil water and salt contents, especially the absolute concentration of soil solution, further affects the photosynthetic efficiency and water use strategy of T. chinensis. Moderate groundwater salinity (brackish and saline groundwater) could improve photosynthetic capacity and reduce WUE of T. chinensis, while excessive high groundwater salinity (salt groundwater) would inhibit photosynthesis severely and improve WUE. T. chinensis has high photosynthetic capacity, wide light ecological amplitude and high WUE under saline groundwater conditions at the groundwater level of 0.9 m. This research can be used for the further study of the relationship between plant photosynthetic processes and soil water and salt contents, and can provide theoretical reference for the restoration and reconstruction of soil and water conservation shelterbelts in the Yellow River Delta.
ZHAO Zi-guo , ZHAO Feng-juan , XIA Jiang-bao , WANG Yue-hai . Effects of groundwater salinities on photosynthesis and water consumption characteristics of Tamarix chinensis in the Yellow River Delta[J]. JOURNAL OF NATURAL RESOURCES, 2019 , 34(12) : 2588 -2600 . DOI: 10.31497/zrzyxb.20191209
Table 1 Soil water and salt parameters under different groundwater mineralization (%)表1 不同地下水矿化度下土壤水盐参数 |
| 地下水矿化度 | 土壤含盐量 | 土壤含水量 | 土壤溶液绝对浓度 |
|---|---|---|---|
| 淡水 | 0.08±0.02a | 18.25±0.92a | 0.46±0.11a |
| 微咸水 | 0.14±0.01b | 19.15±0.94ab | 0.76±0.09a |
| 咸水 | 0.31±0.02c | 20.90±0.95bc | 1.48±0.14b |
| 盐水 | 0.58±0.04d | 21.12±1.22c | 2.75±0.34c |
注:表中数据为平均值±标准误,n=3,同列不同小写字母表示差异显著(P<0.05),下同。 |
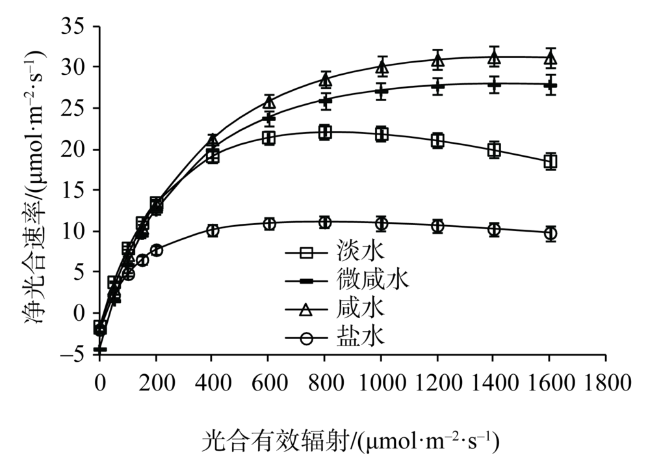
Fig. 1 Light response of net photosynthetic rate in leaves of Tamarix chinensis under different groundwater salinities图1 不同地下水矿化度下柽柳叶片光合光响应 |
Table 2 Photosynthetic parameters of Tamarix chinensis leaves under different groundwater salinities表2 不同地下水矿化度下柽柳叶片光合作用参数 |
| 矿化度 | 最大净光合速率 /(μmol·m-2·s-1) | 光饱和点 /(μmol·m-2·s-1) | 光补偿点 /(μmol·m-2·s-1) | 表观量子效率 /(mol·mol-1) | 暗呼吸速率 /(mmol·m-2·s-1) |
|---|---|---|---|---|---|
| 淡水 | 22.17±1.07c | 832±22a | 35.9±1.4d | 0.0817±0.003b | 4.37±0.18c |
| 微咸水 | 27.92±1.20b | 1439±47b | 13.1±0.6a | 0.0972±0.0038c | 1.58±0.13a |
| 咸水 | 31.30±1.35a | 1462±49b | 16.7±0.7b | 0.0832±0.0027b | 1.71±0.12a |
| 盐水 | 11.14±0.84d | 792±26a | 20.5±0.9c | 0.0579±0.0024a | 2.00±0.15b |
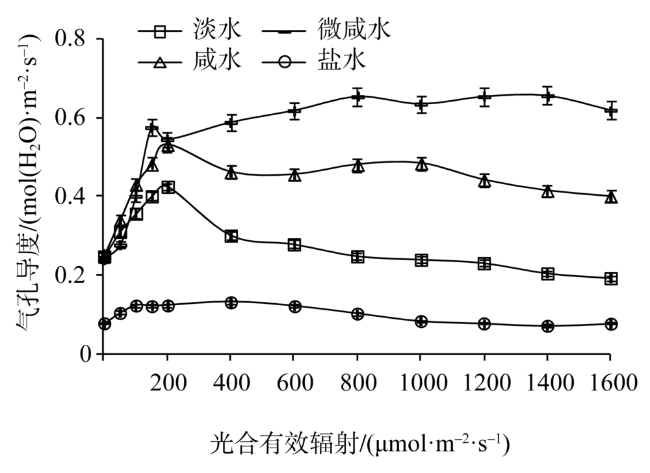
Fig. 2 Light response of stomatal conductance in leaves of Tamarix chinensis under different groundwater salinities图2 不同地下水矿化度下柽柳叶片气孔导度光响应 |
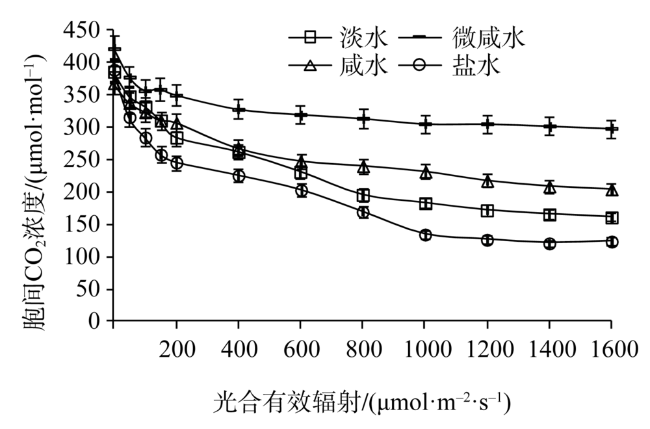
Fig. 3 Light response of intercellular CO2 concentration in leaves of Tamarix chinensis under different groundwater salinities图3 不同地下水矿化度条件下柽柳叶片胞间CO2浓度光响应 |
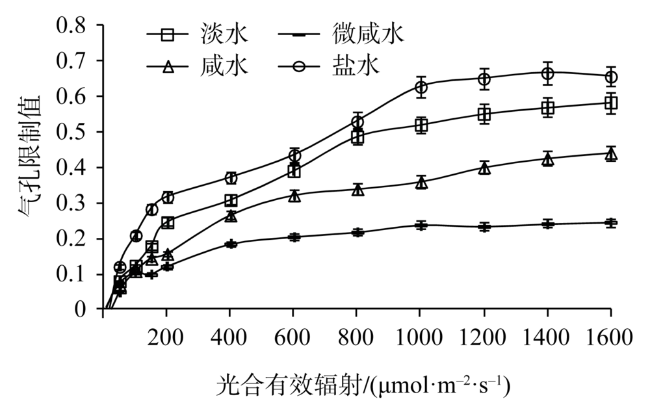
Fig. 4 Light response of stomatal limitation in leaves of Tamarix chinensis under different groundwater salinities图4 不同地下水矿化度下柽柳叶片气孔限制值光响应 |
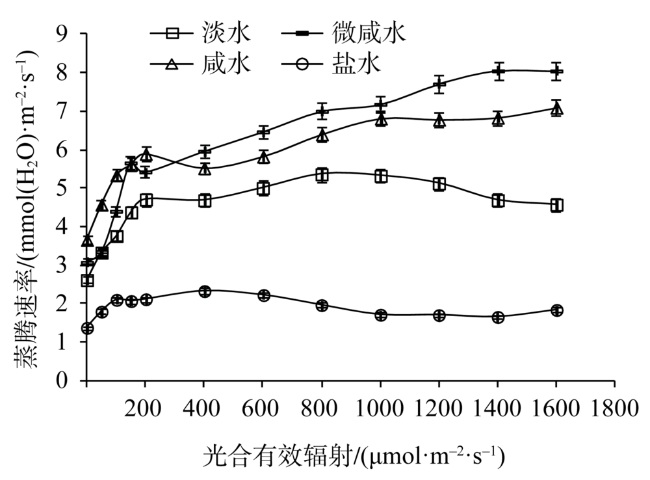
Fig. 5 Light response of transpiration rate in leaves of Tamarix chinensis under different groundwater salinities图5 不同地下水矿化度条件下柽柳叶片蒸腾速率光响应 |
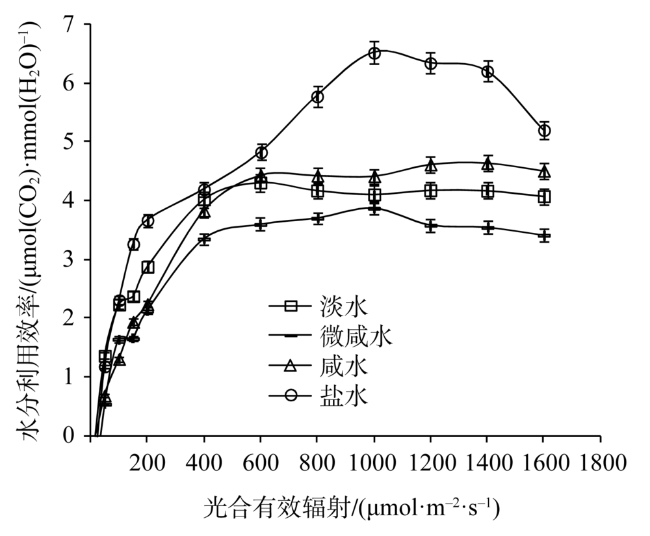
Fig. 6 Light response of water use efficiency in leaves of Tamarix chinensis under different groundwater salinities图6 不同地下水矿化度条件下柽柳叶片水分利用效率光响应 |
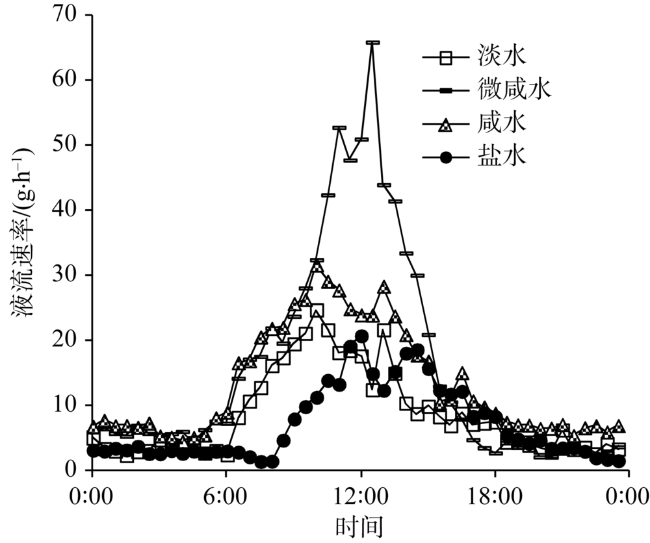
Fig. 7 The daily change of stem sap flow in Tamarix chinensis under different groundwater salinities图7 不同地下水矿化度下柽柳液流速率日变化 |
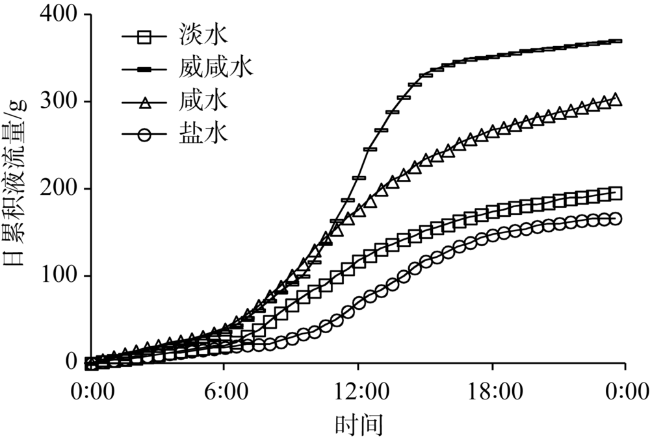
Fig. 8 The daily cumulative sap flow in Tamarix chinensis under different groundwater salinities图8 不同地下水矿化度下柽柳日累积液流量 |
| [1] |
[
|
| [2] |
[
|
| [3] |
[
|
| [4] |
[
|
| [5] |
[
|
| [6] |
[
|
| [7] |
[
|
| [8] |
[
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
[
|
| [13] |
[
|
| [14] |
[
|
| [15] |
[
|
| [16] |
[
|
| [17] |
[
|
| [18] |
[
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
[
|
| [25] |
[
|
| [26] |
[
|
| [27] |
|
| [28] |
[
|
| [29] |
[
|
| [30] |
[
|
| [31] |
GOEDHART,
|
| [32] |
|
| [33] |
[
|
| [34] |
[
|
| [35] |
|
| [36] |
[
|
| [37] |
|
| [38] |
|
| [39] |
[
|
| [40] |
[
|
| [41] |
[
|
| [42] |
[
|
| [43] |
[
|
| [44] |
|
| [45] |
|
| [46] |
[
|
| [47] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}