
自然保护地跨区联合保护可以降低保护资金投入吗?——基于保护生物学的空间效应分析
|
高燕(1980- ),女,湖北荆门人,博士,副教授,研究方向为自然保护地体制建设、生态旅游。E-mail: ygao@hbue.edu.cn |
收稿日期: 2022-03-21
修回日期: 2022-10-08
网络出版日期: 2023-04-11
基金资助
教育部人文社会科学研究规划基金项目(22YJAZH021)
Does cross-regional natural conservation reduce the investment of conservation funds:Spatial effect analysis based on Conservation Biology
Received date: 2022-03-21
Revised date: 2022-10-08
Online published: 2023-04-11
“山川形便”的行政区划原则人为割裂生境,造成生境破碎化,引致负面边缘效应,需要更高成本去维持生物多样性,增加了自然保护财政资金投入。实施跨区联合保护可有效解决上述问题。引入保护生物学边缘效应理论,构建了以跨区联合保护为核心解释变量的自然保护资金投入空间计量理论模型。以31个省份的自然保护区为样本的实证研究结果表明:跨区联合保护是导致自然保护投资水平在邻接行政区之间存在正向空间依赖性的主导因素,且跨区联合保护强度越大,越能降低邻接地区保护资金投入水平;建议在省级和中央层面建立自然保护地联合保护机构与协调机制;全面开启跨行政区保护地在界限范围、保护焦点上的优化整合;联合保护地的管理机构应针对焦点物种设置统筹使用的专项保护资金,以更高效率的资金投入促进多地协同解决生境破碎化问题。
高燕 , 邓毅 . 自然保护地跨区联合保护可以降低保护资金投入吗?——基于保护生物学的空间效应分析[J]. 自然资源学报, 2023 , 38(4) : 1040 -1057 . DOI: 10.31497/zrzyxb.20230414
The administrative division principle of "mountains and rivers form convenience" artificially splits the continuous habitat, leading to habitat fragmentation. Habitat fragmentation has a negative edge effect, which requires a higher cost to maintain biodiversity, resulting in inefficient financial investment in nature conservation. The implementation of cross-regional natural conservation is an effective way to solve the above problems. This study introduces the edge effect theory of conservation biology, traces the theoretical basis for the spatial spillover effect of cross-regional natural conservation on nature conservation investment, and constructs a spatial econometric theoretical model of the factors affecting the level of nature conservation investment. Taking nature reserves in 31 provincial-level regions as samples, the empirical test results of the model show that the investment level of nature conservation has a positive spatial dependence among different administrative regions, and cross-regional natural conservation is the dominant factor leading to the spatial effect of the investment level of nature conservation, and the greater the intensity of cross-regional natural conservation, the lower the investment level of financial funds in contiguous areas. Finally, the study puts forward the following suggestions: establish cross-regional natural conservation institutions and coordination mechanisms for nature reserves at the provincial and national levels, and fully open the optimization and integration of cross administrative protected areas in terms of boundary scope and protection focus. The management organization of joint protected areas should strengthen the overall use of financial funds, set up special protection funds for focus species, and promote the coordinated solution of habitat fragmentation by more efficient financial investment.
表1 中国自然保护区跨省连片概况Table 1 Overview of China's nature reserves in interprovincial contiguous areas |
| 相邻省(市、自治区) | 自然保护区 | u_area/hm2 | 成立时间 | 相邻省(市、自治区) | 自然保护区 | u_area/hm2 | 成立时间 |
|---|---|---|---|---|---|---|---|
| 新疆、甘肃 | 罗布泊野骆驼 | 6120000 | 19860909 | 江西、福建 | 赣江源 | 16100.85 | 19980517 |
| 安南坝野骆驼 | 396000 | 19821119 | 汀江源 | 10379.7 | 20010609 | ||
| 新疆、青海、西藏 | 可可西里 | 4500000 | 19951005 | 江西武夷山 | 16007 | 19810306 | |
| 羌塘 | 29800000 | 19930716 | 福建武夷山 | 56527 | 19790703 | ||
| 阿尔金山 | 4500000 | 19830516 | 安徽、浙江 | 安徽清凉峰 | 7811.2 | 19790101 | |
| 甘肃、青海 | 甘肃祁连山 | 1987200 | 19871024 | 临安清凉峰 | 10800 | 19850807 | |
| 青海祁连山 | 834700 | 20051230 | 湖北、河南、安徽 | 湖北丹江口库区湿地 | 45103 | 20030318 | |
| 甘肃、四川、陕西 | 白水江 | 183799 | 19630101 | 丹江湿地 | 64027 | 20010810 | |
| 王朗 | 32297 | 19630402 | 大别山 | 16048.2 | 20030928 | ||
| 唐家河 | 40000 | 19781215 | 河南大别山 | 10600 | 19820624 | ||
| 九寨沟 | 64297 | 19781215 | 金寨天马 | 28913.7 | 19820601 | ||
| 青木川 | 10200 | 20030521 | 山西、河南 | 阳城莽河猕猴 | 5600 | 19831226 | |
| 甘肃、宁夏 | 太统—崆峒山 | 16283 | 19821119 | 太行山猕猴 | 56600 | 19820624 | |
| 六盘山 | 26784 | 19820701 | 河北、山西 | 驼梁 | 21311.9 | 20010331 | |
| 陕西、甘肃 | 陕西子午岭 | 40621 | 19990209 | 亚高山草甸 | 3333 | 19861201 | |
| 庆阳子午岭 | 242133 | 20051011 | 河北、北京 | 大海陀 | 12634 | 19990707 | |
| 宁夏、内蒙古 | 宁夏贺兰山 | 193535.7 | 19820701 | 松山 | 6212.96 | 19850401 | |
| 内蒙古贺兰山 | 67710.6 | 19920513 | 内蒙古、河北 | 黑里河 | 27638 | 19961231 | |
| 沙坡头 | 14043.09 | 19840901 | 茅荆坝 | 40038 | 20020529 | ||
| 内蒙古腾格里沙漠 | 721707 | 20030301 | 内蒙古、辽宁 | 大黑山 | 86799.4 | 19961022 | |
| 重庆、四川 | 大巴山 | 115750 | 19790501 | 努鲁儿虎山 | 16705.6 | 20001101 | |
| 花萼山 | 48203.39 | 19960101 | 内蒙古、黑 龙江 | 大兴安岭汗马 | 107348 | 19950529 | |
| 重庆、湖北 | 五里坡 | 35276.6 | 20000929 | 呼中 | 167213 | 19840509 | |
| 阴条岭 | 22423.1 | 20000810 | 内蒙古、吉林 | 科尔沁 | 126987 | 19850209 | |
| 神农架 | 70467 | 19820301 | 向海 | 105467 | 19810320 | ||
| 湖北、湖南 | 五峰后河 | 10340 | 19850303 | 黑龙江、吉林 | 老爷岭东北虎 | 71278 | 20111223 |
| 壶瓶山 | 66568 | 19820403 | 珲春东北虎 | 108700 | 20011022 | ||
| 七姊妹山 | 34550 | 19900228 | 汪清 | 67434 | 20021220 | ||
| 八大公山 | 20000 | 19820403 | 甘肃、河南、山东、(黄河) | 黄河首曲 | 203401 | 19951123 | |
| 贵州、广西 | 茂兰 | 20000 | 19840101 | 新乡黄河湿地鸟类 | 22780 | 19880727 | |
| 木论 | 8969 | 19910622 | 河南黄河湿地 | 68000 | 19950818 | ||
| 湖南、广西 | 永州都庞岭 | 20066.4 | 19820101 | 黄河三角洲 | 153000 | 19901227 | |
| 千家洞 | 12231 | 19820608 | 湖北、上海(长江) | 长江天鹅洲白鱀豚 | 2000 | 19901201 | |
| 湖南、广东 | 莽山 | 19833 | 19820403 | 长江新螺段白鱀豚 | 13500 | 19870101 | |
| 南岭 | 50000 | 19840429 | 长江上游珍稀、特有鱼类 | 33174.2 | 19971208 | ||
| 江西、广东 | 桃江源 | 12240 | 20061008 | 九段沙湿地 | 42020 | 20000306 | |
| 九连山 | 13411.6 | 19810306 | 崇明东滩鸟类 | 24155 | 19981116 | ||
| 车八岭 | 7545 | 19810704 | 西藏、云南 | 芒康滇金丝猴 | 185300 | 19930716 | |
| 湖南、江西 | 炎陵桃源洞 | 23786 | 19820403 | 白马雪山 | 276400 | 19830427 | |
| 井冈山 | 21499 | 19810306 |
表2 变量统计性描述Table 2 Descriptive statistics of variables |
| 变量 | 变量说明 | 样本 | 均值 | 标准差 | 最小值 | 最大值 |
|---|---|---|---|---|---|---|
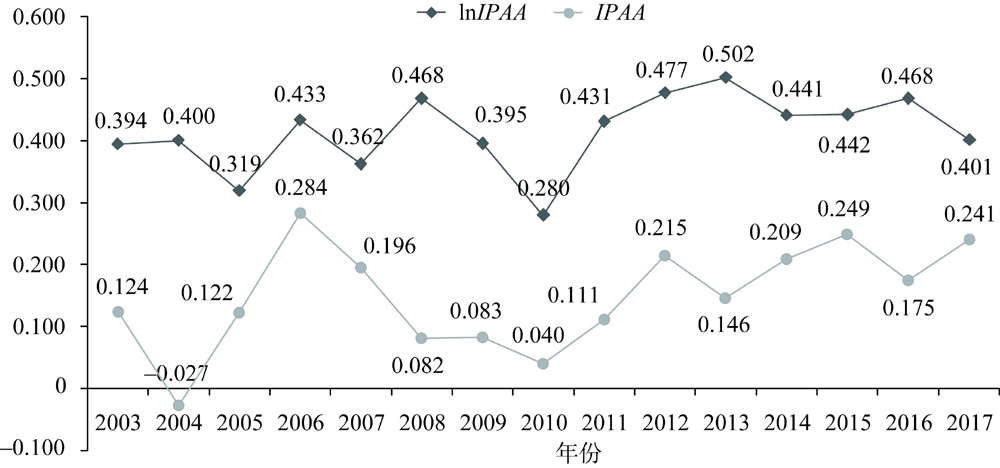
| IPAA | 单位保护面积上自然保护区资金投入/(元/百hm2) | 31×15 | 0.896 | 2.351 | 0.000 | 22.508 |
| lnIPAA | IPAA的对数值 | 31×15 | -1.511 | 1.796 | -11.521 | 3.114 |
| u_area | 与邻省连接的自然保护区面积/万hm2 | 31×15 | 170.182 | 557.670 | 0.000 | 3000.00 |
| APAP | 跨省邻接自然保护区占全省自然保护区面积比例/% | 31×15 | 0.176 | 0.199 | 0.000 | 0.744 |
| APA | 省域自然保护区总面积/万hm2 | 31×15 | 392.055 | 811.526 | 2.300 | 4206.74 |
| TEC | 国内专利申请数/万件 | 31×15 | 4.658 | 8.490 | 0.0024 | 62.783 |
| DPI | 可支配收入/万元 | 31×15 | 1.970 | 1.011 | 0.653 | 6.260 |
| PPD | 人口密度/(人/万km2) | 31×15 | 426.90 | 633.05 | 2.22 | 3850.29 |
| NSP | 单位保护面积上工作人员数量/(人/万hm2) | 31×15 | 4.823 | 6.149 | 0.002 | 28.500 |
| NAP | 国家级自然保护区占全省自然保护区总面积比例/% | 31×15 | 0.387 | 0.280 | 0.009 | 1.000 |
表3 研究变量空间自相关检验结果Table 3 Spatial autocorrelation test results |
| lnIPAA | IPAA | APAP | PPD | DPI | TEC | NSP | NAP | |
|---|---|---|---|---|---|---|---|---|
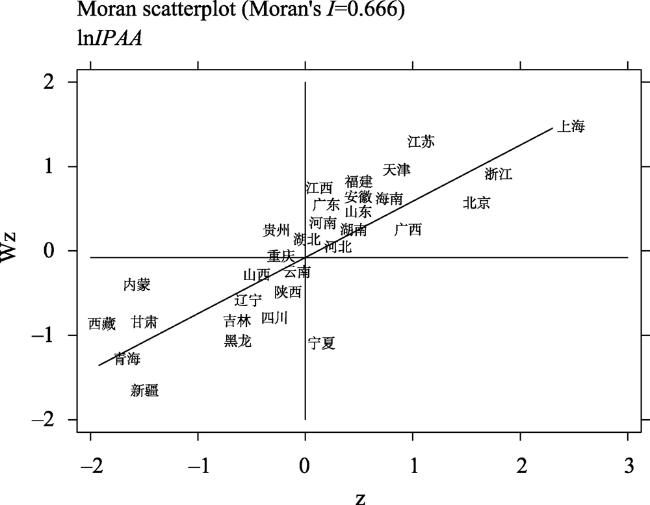
| Moran's I | 0.485*** | 0.214*** | 0.020 | 0.175*** | 0.749*** | 0.348*** | 0.061** | 0.076*** |
| Mean | -0.002 | -0.002 | -0.002 | -0.002 | -0.002 | -0.002 | -0.002 | -0.002 |
| Std dev | 0.031 | 0.029 | 0.030 | 0.030 | 0.031 | 0.030 | 0.030 | 0.031 |
| z-score | 15.956 | 7.383 | 0.746 | 5.886 | 24.545 | 11.602 | 2.080 | 2.557 |
| p-value | 0.000 | 0.000 | 0.228 | 0.000 | 0.000 | 0.000 | 0.019 | 0.005 |
| Geary's C | 0.421*** | 0.377*** | 0.518*** | 0.349*** | 0.116*** | 0.640*** | 1.005 | 0.760*** |
| Mean | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 | 1.000 |
| Std dev | 0.052 | 0.121 | 0.081 | 0.083 | 0.046 | 0.081 | 0.061 | 0.039 |
| z-score | -11.191 | -5.162 | -5.964 | -7.808 | -19.216 | -4.436 | 0.081 | -6.183 |
| p-value | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.468 | 0.000 |
注:*、**、***分别表示p<0.1、p<0.05、p<0.01,下同。 |
表4 模型空间相关性的表现形式判断Table 4 Diagnostic tests for spatial dependence in OLS regression |
| Test | Statistic | p-value |
|---|---|---|
| Spatial error: | ||
| Lagrange multiplier | 16.191 | 0.000 |
| Robust Lagrange multiplier | 0.809 | 0.369 |
| Spatial lag: | ||
| Lagrange multiplier | 46.777 | 0.000 |
| Robust Lagrange multiplier | 31.394 | 0.000 |
表5 空间回归模型LR检验结果Table 5 Likelihood-ratio test |
| SLM nested in SDM | SEM nested in SDM | ||||
|---|---|---|---|---|---|
| LR chi2 | Prob>chi2 | LR chi2 | Prob>chi2 | ||
| 固定效应 | 24.53*** | 0.0004 | 23.51*** | 0.0006 | |
| 随机效应 | 15.32** | 0.0179 | 22.90*** | 0.0008 | |
表6 SDM固定效应模型稳健性估计结果Table 6 Robustness estimation results of SDM fixed effects model |
| 变量 | (1) | (2) | |||||
|---|---|---|---|---|---|---|---|
| 省份固定 | 年份固定 | 双固定 | 省份固定 | 年份固定 | 双固定 | ||
| W×APAP | -0.0915** | -0.0433*** | -0.1160*** | -0.2108*** | -0.0116 | -0.0834** | |
| (-2.4605) | (-4.3179) | (-3.0552) | (-5.8500) | (-0.8264) | (-2.2807) | ||
| 控制变量 | 设置 | 设置 | 设置 | 未设置 | 未设置 | 未设置 | |
| 0.2373*** | 0.0726 | 0.1363** | 0.4839*** | 0.6356*** | 0.1663*** | ||
| (-4.4196) | (-1.3787) | (-2.4442) | (-11.1791) | (-9.1066) | (-3.0856) | ||
| sigma2 | 0.7100*** | 1.0188*** | 0.6700*** | 0.7738*** | 1.6095*** | 0.7067*** | |
| (-4.7218) | (-4.6661) | (-4.9152) | (-5.1107) | (-3.9234) | (-5.0037) | ||
| N/个 | 465 | 465 | 465 | 465 | 465 | 465 | |
| R2 | 0.3535 | 0.2741 | 0.3102 | 0.1394 | 0.1468 | 0.0595 | |
| LogL | -583.312 | -664.4135 | -567.6568 | -614.5997 | -797.5268 | -580.5178 | |
表7 自然保护资金投入空间杜宾模型空间效应分解结果Table 7 The spatial Durbin model space effect decomposition results |
| 模型 | 解释变量 | 直接效应 | 间接效应 | 总效应 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 省份固定 | 年份固定 | 双固定 | 省份固定 | 年份固定 | 双固定 | 省份固定 | 年份固定 | 双固定 | ||||
| (1) | APAP | 0.0328 | -0.0108* | 0.0190 | -0.1041** | -0.0464*** | -0.1264*** | -0.0713 | -0.0572*** | -0.1074* | ||
| (1.2640) | (-1.7024) | (0.7313) | (-2.2024) | (-4.5674) | (-2.7455) | (-1.2469) | (-4.8695) | (-1.8624) | ||||
| PPD | 0.0004 | 0.0012* | 0.0026* | -0.0037 | 0.0038*** | 0.0029 | -0.0033 | 0.0050*** | 0.0055 | |||
| (0.5330) | (1.7196) | (1.8892) | (-1.1721) | (4.8330) | (0.8173) | (-1.1401) | (5.8992) | (1.3476) | ||||
| DPI | 0.0001** | -0.0000 | -0.0000 | 0.0000 | -0.0001* | -0.0003*** | 0.0001*** | -0.0001 | -0.0003** | |||
| (2.2089) | (-0.3359) | (-0.9178) | (0.2869) | (-1.6805) | (-2.7553) | (2.6022) | (-1.5996) | (-2.3475) | ||||
| TEC | 0.0000 | 0.0000* | 0.0000* | -0.0000 | 0.0000** | 0.0000 | 0.0000 | 0.0000** | 0.0000** | |||
| (1.0757) | (1.8919) | (1.8653) | (-0.0587) | (2.0805) | (1.6224) | (0.3244) | (2.3871) | (2.3929) | ||||
| NSP | 0.0097 | 0.0533*** | 0.0063 | -0.0003 | -0.0619** | -0.0081 | 0.0093 | -0.0086 | -0.0019 | |||
| (0.8209) | (2.5843) | (0.5030) | (-0.0120) | (-2.0202) | (-0.3123) | (0.3128) | (-0.2511) | (-0.0691) | ||||
| NAP | 0.4697 | 0.4734 | 0.0279 | 0.6485 | 2.1764** | -0.3064 | 1.1183 | 2.6497** | -0.2785 | |||
| (0.9080) | (0.8028) | (0.0410) | (0.5108) | (2.1093) | (-0.2264) | (0.7554) | (2.1923) | (-0.1580) | ||||
| (2) | APAP | -0.0122 | 0.0011 | 0.0360 | -0.3688*** | -0.0231 | -0.0922** | -0.3810*** | -0.0221 | -0.0562 | ||
| (-0.4357) | (0.2009) | (1.2124) | (-5.3267) | (-0.6617) | (-2.2096) | (-4.2693) | (-0.5681) | (-1.0599) | ||||
| [1] |
杨喆, 吴健. 中国自然保护区的保护成本及其区域分布. 自然资源学报, 2019, 34(4): 839-852.
[
|
| [2] |
李俊生, 朱彦鹏. 国家公园资金保障机制探讨. 环境保护, 2015, 43(14): 38-40.
[
|
| [3] |
刘家顺, 张坤, 繆光平. 中国自然保护区投资机制研究. 林业经济, 2000, (3): 12-17.
[
|
| [4] |
[
|
| [5] |
武晶, 刘志民. 生境破碎化对生物多样性的影响研究综述. 生态学杂志, 2014, 33(7): 1946-1952.
[
|
| [6] |
|
| [7] |
赵鑫蕊, 何思源, 苏杨. 生态系统完整性在管理层面的体现方式: 以跨省国家公园统一管理的体制机制为例. 生物多样性, 2022, 30(3): 178-185.
[
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
李晓肃, 邓武功, 李泽, 等. 自然保护地整合优化: 思路、应对与探讨. 中国园林, 2020, 36(11): 25-28.
[
|
| [17] |
|
| [18] |
余付勤, 张百平, 王晶, 等. 国外大尺度生态廊道保护进展与秦岭国家公园建设. 自然资源学报, 2021, 36(10): 2478-2490.
[
|
| [19] |
|
| [20] |
代陆娇, 王浩瀚, 徐会明, 等. 中国跨区保护现状分析及发展建议. 大理大学学报, 2020, 5(6): 54-60.
[
|
| [21] |
|
| [22] |
|
| [23] |
赵帅, 何爱平, 彭硕毅. 黄河流域环境规制、区域污染转移与技术创新的空间效应. 经济经纬, 2021, 38(5): 12-21.
[
|
| [24] |
邵帅, 李欣, 曹建华, 等. 中国雾霾污染治理的经济政策选择: 基于空间溢出效应的视角. 经济研究, 2016, 51(9): 73-88.
[
|
| [25] |
张廷海, 王点. 工业集聚、空间溢出效应与地区增长差异: 基于空间杜宾模型的实证分析. 经济经纬, 2018, 35(1): 86-91.
[
|
| [26] |
王华春, 平易, 崔伟. 地方政府环境保护支出竞争的空间效应研究. 广东财经大学学报, 2019, 34(4): 51-61.
[
|
| [27] |
|
| [28] |
樊轶侠, 覃凤琴, 王正早. 中国国家公园资金保障机制研究. 财政科学, 2021, 69(9): 91-98.
[
|
| [29] |
王晓霞, 吴健. 中国自然保护区财政资金投入水平分析. 环境保护, 2017, 45(11): 53-57.
[
|
| [30] |
钱者东, 郭辰, 吴儒华, 等. 中国自然保护区经济投入特征与问题分析. 生态与农村环境学报, 2016, 32(1): 35-40.
[
|
| [31] |
任艳梅. 基于典型区域自然保护区资金投入的绩效评价研究. 北京: 北京林业大学, 2016.
[
|
| [32] |
张书杰, 庄优波. 中国自然保护区现状人类利用强度量化特征分析与管理分类建议. 中国园林, 2020, 36(8): 14-19.
[
|
| [33] |
吴成亮, 周莉, 李强. 中国经济增长与自然保护区建设的实证研究. 北京林业大学学报: 社会科学版, 2009, 8(4): 40-42.
[
|
| [34] |
王昌海, 温亚利, 李强, 等. 秦岭自然保护区群保护成本计量研究. 中国人口·资源与环境, 2012, 22(3): 130-136.
[
|
| [35] |
|
| [36] |
|
| [37] |
刘某承, 王佳然, 刘伟玮, 等. 国家公园生态保护补偿的政策框架及其关键技术. 生态学报, 2019, 39(4): 1330-1337.
[
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
李兰兰, 於冉, 於忠祥, 等. 基于STIRPAT模型的长江经济带农业碳排放时空特征及影响因素分析. 安徽农业大学学报: 社会科学版, 2020, 29(5): 32-37.
[
|
| [42] |
沈坤荣, 李剑. 企业间技术外溢的测度. 经济研究, 2009, 44(4): 77-89.
[
|
| [43] |
陶锋, 赵锦瑜, 周浩. 环境规制实现了绿色技术创新的“增量提质”吗: 来自环保目标责任制的证据. 中国工业经济, 2021, (2): 136-154.
[
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
陈强. 高级计量经济学及Stata应用. 北京: 高等教育出版社, 2014: 575-593.
[
|
| [48] |
|
| [49] |
李婧, 谭清美, 白俊红. 中国区域创新生产的空间计量分析: 静态与动态空间面板模型的实证研究. 管理世界, 2010, (7): 43-55, 65.
[
|
| [50] |
姜磊. 空间回归模型选择的反思. 统计与信息论坛, 2016, 31(10): 10-16.
[
|
| [51] |
翁爱芳, 范少贞, 卢远清, 等. 基于成本效益评估的县域自然保护地优化整合方案比较研究: 以福建省泰宁县为例. 自然资源学报, 2021, 36(8): 2020-2037.
[
|
| [52] |
陈叙图, 魏钰. 中国国家公园生态系统完整性的问题与改进对策. 环境保护, 2019, 47(5): 55-58.
[
|
| [53] |
徐会明, 雷建东, 张志云, 等. 龙马山黑白仰鼻猴跨区域联合保护机制及其初步成效. 林业调查规划, 2018, 43(2): 112-116.
[
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}