
玉米—大豆轮作体系对黑土土壤固氮菌群落结构及其质量的影响
|
冯雪婉(1998- ),女,内蒙古包头人,硕士,研究方向为土壤微生物生态。E-mail: 1156357193@qq.com |
收稿日期: 2021-11-01
修回日期: 2022-04-07
网络出版日期: 2022-12-28
基金资助
吉林省教育厅科学技术研究项目(JJKH20210355KJ)
中央引导地方科技发展资金吉林省基础研究专项(202002013JC)
Effects of corn-soybean rotation system on soil nitrogen-fixing bacteria community structure and quality in black soil
Received date: 2021-11-01
Revised date: 2022-04-07
Online published: 2022-12-28
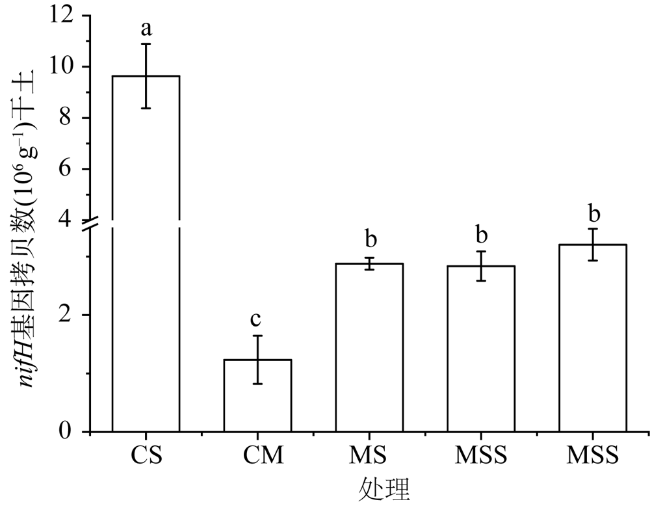
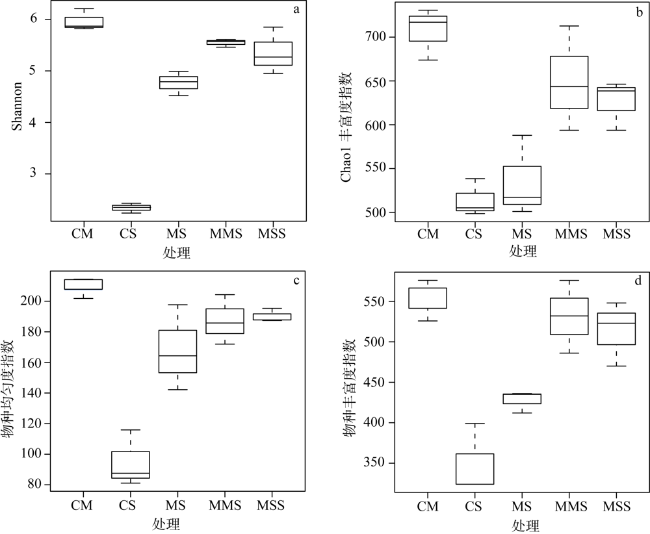
以大豆连作(CS)、玉米连作(CM)、玉米—大豆轮作(MS)、玉米—玉米—大豆(MMS)和玉米—大豆—大豆(MSS)为研究对象,利用荧光定量PCR和高通量测序分析种植制度对土壤固氮菌丰度和群落结构的影响。结果表明:轮作中土壤有机质(SOM)、全磷(TP)、有效氮(AN)和有效磷(AP)的含量显著高于连作;轮作固氮菌丰度显著高于CM,显著低于CS;MMS与MSS固氮菌多样性显著高于CM;轮作和连作土壤固氮菌群落结构差异明显,全氮(TN)是固氮菌群落结构变化的主要驱动因子,种植制度通过土壤化学性质间接影响固氮菌丰度和多样性。这说明,在吉林省西部半干旱区,MSS与MMS更有利于土壤固氮菌繁殖,可以从微生物学的角度为合理种植和氮素调控提供科学依据。
冯雪婉 , 李翠兰 , 彭畅 , 张晋京 , 林园 , 刘杭 . 玉米—大豆轮作体系对黑土土壤固氮菌群落结构及其质量的影响[J]. 自然资源学报, 2022 , 37(9) : 2319 -2333 . DOI: 10.31497/zrzyxb.20220909
The effects of soybean continuous cropping (CS), maize continuous cropping (CM), corn-soybean rotation (MS), corn-corn-soybean (MMS) and corn-soy-soybean (MSS) on soil nitrogen fixing bacteria abundance and community structure were analyzed by fluorescence quantitative PCR and high-throughput sequencing. The results showed that soil organic matter (SOM), total phosphorus (TP), available nitrogen (AN) and available phosphorus (AP) contents in rotation were significantly higher than those in continuous cropping. The abundance of nitrogen-fixing bacteria in rotation was significantly higher than that in CM and significantly lower than that in CS. The diversity of NITR Ogen-fixing bacteria in MMS and MSS was significantly higher than that in CM. Total nitrogen (TN) was the main driving factor for the change of nitrogen-fixing bacteria community structure. Planting system indirectly affected the abundance and diversity of nitrogen-fixing bacteria through soil chemical properties. These results indicate that MSS and MMS are more conducive to the propagation of soil nitrogen-fixing bacteria in the semi-arid area of Western Jilin province, which can provide scientific basis for rational planting and nitrogen regulation from the perspective of microbiology.
表1 土壤化学性质的测定方法Table 1 Determination methods of soil chemical properties |
| 化学性质 | 测定方法 |
|---|---|
| 土壤有机质SOM | K2CrO7氧化—外加热法 |
| pH | pH计土壤pH (水土比为2.5∶1) |
| 土壤全氮TN | 凯氏定氮法 |
| 土壤全磷TP | HClO4-H2SO4消解—钼锑抗比色法 |
| 土壤全钾TK | 火焰分光光度计法 |
| 土壤速效氮AN | 碱解扩散法 |
| 土壤速效磷AP | 钼锑抗比色法 |
| 土壤速效钾AK | 火焰分光光度计法 |
表2 荧光定量PCR的反应体系Table 2 Reaction system of fluorescence quantitative PCR |
| 成分 | 体积/μL |
|---|---|
| nifH-F (10 μM) | 1 |
| nifH-R (10 μM) | 1 |
| SYBR®PremixEx Taq™II (TliRNaseH Plus), ROX plus | 1 |
| 2×Taq Master Mix | 25 |
| 水 | 22 |
| 总共 | 50 |
表3 荧光定量PCR的扩增条件Table 3 Amplification conditions of fluorescence quantitative PCR |
| 程序 | 温度/℃ | 时间/秒 |
|---|---|---|
| 预变性 | 94 | 300 |
| 变性 | 55 | 30 |
| 退火 | 2 | 30 |
| 延伸 | 72 | 600 |
表4 PCR的反应体系Table 4 Reaction systems of high-throughput sequencing |
| 成分 | 体积/μL |
|---|---|
| 2×Taq Plus Master Mix | 12.5 |
| BSA (2 ng/μl) | 3 |
| Forward Primer (5 μM) | 1 |
| Reverse Primer (5 μM) | 1 |
| DNA (加入的DNA总量为30 ng) | 2 |
| dd H2O | 5.5 |
| 总共 | 25 |
表5 PCR的反应参数Table 5 PCR reaction parameters |
| 程序 | 温度/℃ | 时间/秒 |
|---|---|---|
| 预变性 | 95 | 300 |
| 变性 | 55 | 45 |
| 退火 | 72 | 50 |
表6 玉米—大豆轮作和连作下土壤的化学性质Table 6 Soil chemical properties under corn-soybean rotation and continuous cropping |
| 处理 | pH | SOM /(g/kg) | TN /(g/kg) | TP /(g/kg) | TK /(g/kg) | AN /(mg/kg) | AP /(mg/kg) | AK /(mg/kg) |
|---|---|---|---|---|---|---|---|---|
| CS | 6.97±0.06ab | 11.95±1.24c | 1.87±0.01b | 0.55±0..02c | 22.53±0.31c | 109.27±8.48b | 22.28.±0.39b | 156.17±3.01a |
| CM | 6.90±0.09b | 12.99±0.18c | 1.80±0.04c | 0.56±0.02c | 25.53±1.03a | 117.08±2.42b | 22.28±2.68b | 163.00±4.82a |
| MS | 7.15±0.22ab | 15.20±0.15b | 1.88±0.01b | 0.68±0.04a | 24.93±1.45ab | 132.03±11a | 44.13±7.69a | 162.50±15.52a |
| MMS | 7.22±0.15a | 16.83±0.48a | 2.06±0.01a | 0.65±0.05ab | 24.73±0.23ab | 123.67±0.67ab | 36.62±2.61a | 168.33±5.97a |
| MSS | 7.20±0.18a | 17.73±0.82a | 2.09±0.03a | 0.58±0.06bc | 23.60±1.20bc | 123.09±4.03ab | 36.62±8.97a | 159.50±1.80a |
注:同列不同小写字母表示不同处理间差异显著,下同。 |
表7 土壤固氮菌nifH基因 Illumina MiSeq 高通量测序数据(97%相似度)Table 7 Illumina MiSeq sequenced bacterial data (at 97% sequence similarity) based on the nifH gene |
| 处理 | 序列/条 | Final TAG OTU/个 |
|---|---|---|
| CS1 | 142036 | 499 |
| CS2 | 90760 | 324 |
| CS3 | 106553 | 324 |
| CM1 | 141014 | 576 |
| CM2 | 71625 | 526 |
| CM3 | 135340 | 557 |
| MS1 | 26825 | 412 |
| MS2 | 76579 | 436 |
| MS3 | 155086 | 535 |
| MMS1 | 142022 | 532 |
| MMS2 | 142081 | 486 |
| MMS3 | 73582 | 576 |
| MSS1 | 143927 | 523 |
| MSS2 | 168971 | 548 |
| MSS3 | 31076 | 470 |
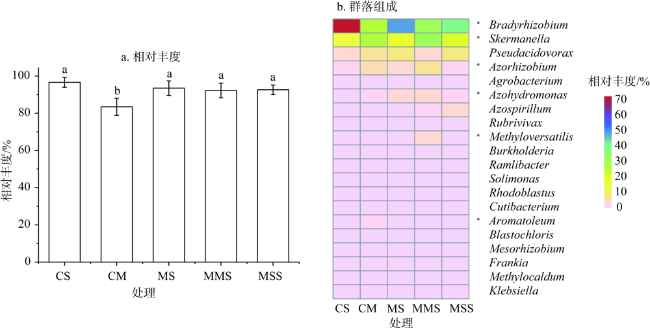
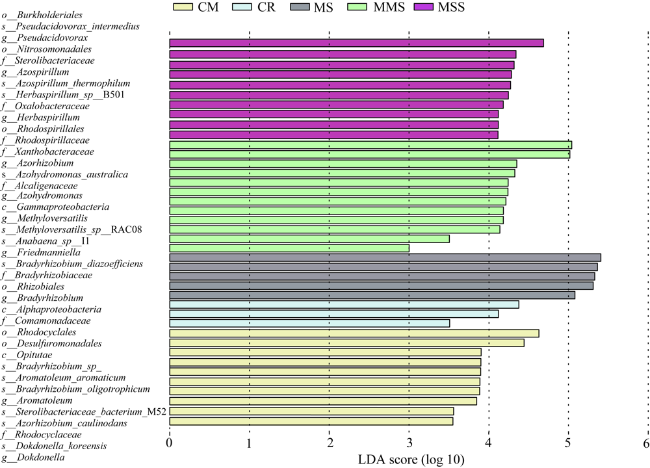
图2 玉米—大豆轮作和连作条件下Proteobacteria的相对丰度和土壤固氮菌优势菌属水平上的群落组成注:*:P<0.05,ANOVA差异分析。颜色代表属相对丰度高低,粉色为低丰度,红色为高丰度。 Fig. 2 The relative abundance of Proteobacteria and the community composition at the level of the dominant genus of soil nitrogen-fixation bacteria under the conditions of corn-soybean rotation and continuous cropping |
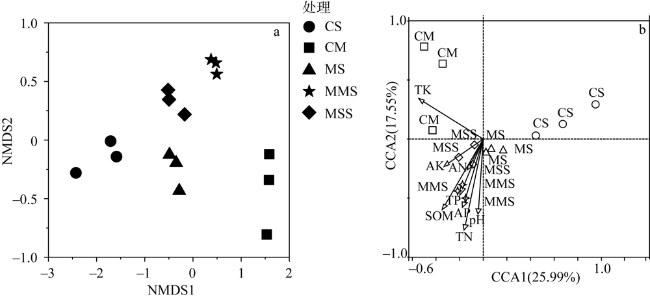
图4 玉米—大豆轮作和连作条件下土壤固氮菌群落的包作非度量多维尺度分析及土壤固氮菌群落结构和环境因子的典范对应分析Fig. 4 Inclusion nonmetric multidimensional scale analysis (NMDS) of soil nitrogen-fixing bacteria community and canonical correspondence analysis (CCA) of soil nitrogen-fixing bacteria community structure and environmental factors under corn-soybean rotation and continuous cropping |
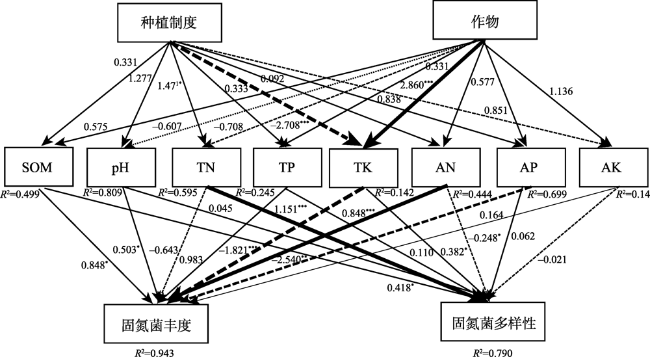
表8 玉米—大豆轮作和连作条件下土壤固氮菌群落结构和环境因子的典范对应分析的解释率和贡献率Table 8 Explanation rate and contribution rate of canonical correspondence analysis of soil nitrogen-fixing bacteria community structure and environmental factors under corn-soybean rotation and continuous cropping |
| 指标 | 解释率/% | 贡献率/% | P |
|---|---|---|---|
| SOM | 4.1 | 6.6 | 0.764 |
| pH | 7.3 | 11.6 | 0.236 |
| TN | 12.4 | 19.6 | 0.01 |
| TP | 5.9 | 9.4 | 0.554 |
| TK | 11.5 | 18.3 | — |
| AN | 7.6 | 12.6 | 0.16 |
| AP | 7.9 | 12.6 | 0.146 |
| AK | 5.9 | 9.3 | — |
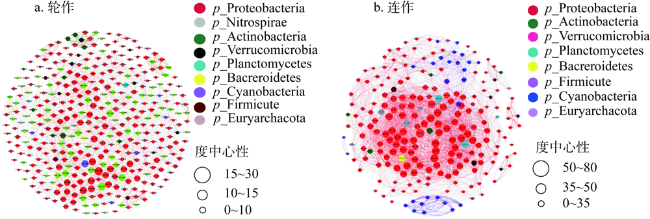
图7 玉米—大豆轮作和连作土壤固氮菌共线性网络分析注:节点颜色表示不同土壤固氮菌门,氮菌网络图红线表示两个节点之间的正相关,绿线表示负相关。 Fig. 7 Soil consolidation in corn-soybean rotation and continuous cropping |
表9 土壤固氮菌网络的拓扑特征Table 9 Topological characteristics of nitrogen-fixing bacteria network |
| 处理 | 连作 | 轮作 |
|---|---|---|
| 节点/个 | 224 | 296 |
| 边/条 | 2530 | 693 |
| 平均度/(条/个) | 28.657 | 6.037 |
| 网络直径/链路树 | 6 | 12 |
| 平均路径长度/链路树 | 2.682 | 4.719 |
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
翁佩莹, 郑红艳. 作物连作障碍的成因与机制及其消减策略. 亚热带植物科学, 2020, 49(2): 157-162.
[
|
| [5] |
梁鑫源, 金晓斌, 孙瑞, 等. 粮食安全视角下的土地资源优化配置及其关键问题. 自然资源学报, 2021, 36(12): 3031-3053.
[
|
| [6] |
林敏. 加强生物固氮科技创新, 支撑“藏粮于地”国家战略. 农经, 2021, (3): 51-53.
[
|
| [7] |
关大伟, 李力, 岳现录, 等. 我国大豆的生物固氮潜力研究. 植物营养与肥料学报, 2014, 20(6): 1497-1504.
[
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
韩博, 金晓斌, 顾铮鸣, 等. 乡村振兴目标下的国土整治研究进展及关键问题. 自然资源学报, 2021, 36(12): 3007-3030.
[
|
| [14] |
|
| [15] |
鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 1999: 146-296.
[
|
| [16] |
ROSCH,
|
| [17] |
何冬华, 陈俊辉, 徐秋芳, 等. 集约经营对毛竹林土壤固氮细菌群落结构和丰度的影响. 应用生态学报, 2015, 26(10): 52-59.
[
|
| [18] |
|
| [19] |
曲晓华, 赵晓燕, 马杰, 等. 大豆根系分泌中特定物质对土壤微生物活性的影响. 福建农林学报, 2015, 30(3): 298-302.
[
|
| [20] |
刘苹, 赵海军, 仲子文, 等. 三种根系分泌脂肪酸对花生生长和土壤酶活性的影响. 生态学报, 2013, 33(11): 3332-3339.
[
|
| [21] |
|
| [22] |
严君, 韩晓增, 陈旭, 等. 施肥对小麦、玉米和大豆连作土壤微生物群落功能多样性的影响. 干旱地区农业研究, 2019, 37(6): 171-177.
[
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
姜南. 茎瘤固氮根瘤菌(Azorhizobium caulinodans ORS571) 可溶性趋化受体的功能研究. 北京: 中国科学院大学.
[
|
| [37] |
王磊, 王静, 张爱君, 等. 小麦—甘薯轮作长期增施有机肥对碱性土壤固氮菌群落结构及多样性的影响. 生态学报, 2020, 40(16): 5771-5782.
[
|
| [38] |
王光华, 金剑, 刘晓冰. 农田分子生态学科组10年来研究进展回顾及展望. 土壤与作物, 2018, 7(2): 212-221.
[
|
| [39] |
|
| [40] |
周岚, 杨永, 王海占, 等. 玉米—大豆轮作及氮肥施用对土壤细菌群落结构的影响. 作物学报, 2013, 39(11): 2016-2022.
[
|
| [41] |
丁俊男, 王慧, 于少鹏, 等. 玉米—大豆轮作对土壤酶和根系微生物群落多样性的影响. 黑龙江农业科学, 2021, (4): 11-16.
[
|
| [42] |
董艳, 汤利, 郑毅, 等. 施氮对间作蚕豆根际微生物区系和枯萎病发生的影响. 生态学报, 2010, 3(7): 1797-1850.
[
|
| [43] |
|
| [44] |
INDERJIT. Soil microorganisms: An important determinant of allelopathic activity. Plant and Soil, 2005, 274: 227-236.
|
| [45] |
蔡树美, 徐四新, 张翰林, 等. 滩涂土壤固氮菌群落与环境因子的典范对应分析. 土壤, 2017, 49(6): 1159-1196.
[
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
汪峰,
[
|
| [52] |
|
| [53] |
|
/
| 〈 |
|
〉 |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}