
全球变化背景下南方红豆杉地域分布变化
|
李艳红(1994- ),女,河南信阳人,硕士,主要从事气候变化和山地资源研究。E-mail: lyh6299@126.com |
收稿日期: 2019-07-12
要求修回日期: 2019-12-05
网络出版日期: 2021-05-28
基金资助
国家自然科学基金项目(41671090)
科技基础资源调查专项(2017FY100902)
版权
Changes of Taxus chinensis var. mairei habitat distribution under global climate change
Received date: 2019-07-12
Request revised date: 2019-12-05
Online published: 2021-05-28
Copyright
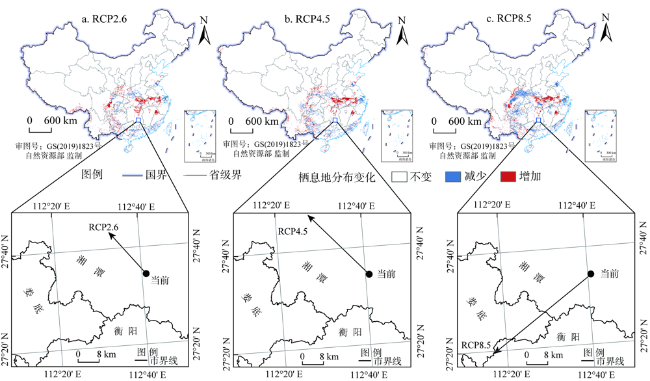
气候是影响植物栖息地的重要因素之一,预测气候变化对植物潜在分布范围变动的影响,对促进植物资源的可持续利用具有重要意义。基于最大熵Maxent模型结合11个环境变量,预测2050s三种气候情景下(RCP2.6、RCP4.5和RCP8.5)南方红豆杉(Taxus chinensis var. mairei)在中国的潜在地理分布状况,分析影响其分布的主要因素,探讨其分布格局的改变对我国亚热带北界的指示意义。结果表明:(1)南方红豆杉的适宜栖息地(生境指数P >0.2)主要分布在我国亚热带暖温带季风区,绝大部分核心栖息地(生境指数P >0.6)分布在秦岭大巴山以南地区;(2)Jackknife测试结果显示,最冷季降水量(bio19)、气温平均日较差(bio2)、气温年变化范围(bio7)、最暖季平均温度(bio10)和海拔(Elev)对南方红豆杉空间分布适宜性影响最大; (3)随气候变化,2050s南方红豆杉有沿纬度向北和海拔向上迁移的趋势,并且我国亚热带北界受气候变化的影响将逐渐向北移动。
李艳红 , 张立娟 , 朱文博 , 张静静 , 徐帅博 , 朱连奇 . 全球变化背景下南方红豆杉地域分布变化[J]. 自然资源学报, 2021 , 36(3) : 783 -792 . DOI: 10.31497/zrzyxb.20210318
Climate is an important factor affecting plant habitat. It is of great significance to promote the sustainable use of natural resources and predict the impact of climate change on the potential distribution of plants. The potential geographical distribution of Taxus chinensis var. mairei in China was simulated by the maximum entropy model (Maxent), based on 11 environmental variables under three climate scenarios (RCP2.6, RCP4.5 and RCP8.5) in the 2050s. And we discussed the significance of its distribution patterns to the northern boundary of subtropical China. The results showed that AUC=0.950, indicating that Maxent had high accuracy and reliability in prediction of the potential distribution for Taxus chinensis var. mairei. Jackknife test results showed that the main environmental variables affecting the distribution suitability of Taxus chinensis var. mairei. were the precipitation of coldest quarter (bio19), mean diurnal range (bio2), temperature annual range (bio7), mean temperature of warmest quarter (bio10) and altitude (Elev), and the cumulative contribution of >90%. The suitable habitats (P>0.2) of Taxus chinensis var. mairei are mainly distributed in the subtropical and warm temperate monsoon areas of 18°-36°N and 104°-124°E in China, and most of the core habitats (P>0.6) were distributed to the south of Qinling-Daba Mountains. With the increase of greenhouse gas concentrations in the 2050s, the suitable habitats of Taxus chinensis var. mairei increased in RCP2.6 scenario, and decreased in RCP4.5 and RCP8.5 scenarios. With the climate warming in the future, the suitable habitats range of Taxus chinensis var. mairei in RCP2.6 and RCP4.5 scenarios are expanding north- and westward, while shrinking south- and eastward. In the RCP8.5 scenario, the suitable habitats distribution of Taxus chinensis var. mairei are "expanding westward", and "shrinking eastward". With climate change, the centroid of Taxus chinensis var. mairei distribution moved to the northwest under the scenarios of RCP2.6 and RCP4.5, and moved to the southwest in the RCP 8.5 scenario. The distribution of Taxus chinensis var. mairei presents a rising trend as the altitude and the latitude increase in the 2050s, but the migration rate is low and the spread range is small. And the northern boundary of subtropical China will move northward under climate change.
表1 南方红豆杉环境变量表Table 1 Environmental variables of Taxus chinensis var. mairei |
| 环境变量 | 描述 | 贡献率/% | 最适范围 |
|---|---|---|---|
| bio2 | 气温平均日较差 | 21.2 | 0~11 ℃ |
| bio3 | 等温性 | 3.2 | >25% |
| bio6 | 最冷月最低温度 | 1.3 | <5 ℃ |
| bio7 | 气温年变化范围 | 8.1 | 1~16 ℃ |
| bio10 | 最暖季平均温度 | 5.5 | 3~22 ℃ |
| bio13 | 最湿月降水量 | 1.1 | >155 mm |
| bio15 | 降水季节性变化 | 0.2 | 20%~70% |
| bio19 | 最冷季降水量 | 52.8 | >25 mm |
| T_BS | 表层土壤基本饱和度 | 2.7 | 4%~60% |
| AWC_CLASS | 土壤有效含水量 | 0.6 | 7.5%~15% |
| Elev | 海拔 | 3.3 | >0 m |
表2 南方红豆杉适宜栖息地(P >0.2)分布面积Table 2 Suitable habitats (P >0.2) of Taxus chinensis var. mairei (104 km2) |
| 生境指数P | 当前(1960—1990年) | 未来(2050s) | ||
|---|---|---|---|---|
| RCP2.6 | RCP4.5 | RCP8.5 | ||
| 0.2~0.4 | 57.50 | 60.48 | 61.82 | 68.56 |
| 0.4~0.6 | 37.89 | 40.22 | 42.40 | 37.24 |
| >0.6 | 52.74 | 49.01 | 43.46 | 31.79 |
| 合计(>0.2) | 148.13 | 149.71 | 147.68 | 137.59 |
| [1] |
IPCC. Climate Change 2014:The physical science basis. Contribution of Working Group I to the fifth Assessment Report of the Intergovernmental Panel on Climate Change. New York: Cambridge University Press, 2014.
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
邓晨晖, 白红英, 高山, 等. 秦岭植被覆盖时空变化及其对气候变化与人类活动的双重响应. 自然资源学报, 2018,33(3):425-438.
[
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
包维楷, 陈庆恒. 中国的红豆杉资源及其开发研究现状与发展对策. 自然资源学报, 1998,13(4):375-380.
[
|
| [15] |
廖文波, 张志权, 苏志尧. 抗癌植物南方红豆杉保护生物学价值的评价. 生态科学, 1996,15(2):17-20.
[
|
| [16] |
高润梅, 石晓东, 樊兰英, 等. 山西省南方红豆杉自然分布与群落生态学特征. 应用生态学报, 2016,27(6):1820-1828.
[
|
| [17] |
罗宁, 欧建德, 吴志庄. 福建南方红豆杉林生境质量评价研究. 西南林业大学学报, 2016,36(6):52-57.
[
|
| [18] |
朱志军. 南方红豆杉造林关键技术研究. 杭州: 浙江农林大学, 2011.
[
|
| [19] |
王艺, 张蕊, 冯建国. 不同种源南方红豆杉生长差异分析及早期速生优良种源筛选. 植物资源与环境学报, 2012,21(4):41-47.
[
|
| [20] |
吴普, 王丽丽, 黄磊. 五个中国特有针叶树种树轮宽度对气候变化的敏感性. 地理研究, 2006,25(1):43-52.
[
|
| [21] |
陈涵, 张超, 余树全. 基于Stacking模型集成算法的莲都区南方红豆杉潜在分布区. 浙江农林大学学报, 2019,36(3):69-75.
[
|
| [22] |
谢伟东. 南岭山地南方红豆杉遗传变异与气候因子相关性研究. 长沙: 中南林业科技大学, 2017.
[
|
| [23] |
程蓓蓓. 中国红豆杉属分子谱系地理学与遗传多样性研究. 北京: 中国林业科学研究院, 2016.
[
|
| [24] |
付晓峰, 张桂萍, 吴伟, 等. 不同产地南方红豆杉根际AMF多样性. 菌物学报, 2017,36(8):1061-1071.
[
|
| [25] |
张雪梅, 李德铢, 高连明. 南方红豆杉谱系地理学研究. 西北植物学报, 2012,32(10):1983-1989.
[
|
| [26] |
|
| [27] |
茹文明. 濒危植物南方红豆杉生态学研究. 太原: 山西大学, 2006.
[
|
| [28] |
|
| [29] |
|
| [30] |
张静静, 王岩松, 朱连奇, 等. 近50年来豫西山地亚热带北界变化分析. 河南大学学报: 自然科学版, 2016,46(1):40-49.
[
|
| [31] |
杨强, 郑西楠, 何立恒. 基于HadCM3模式的我国主要气候区划界线时空预测研究. 干旱区地理, 2017,40(1):17-25.
[
|
| [32] |
莫兴国, 胡实, 卢洪健, 等. GCM预测情景下中国21世纪干旱演变趋势分析. 自然资源学报, 2018,33(7):1244-1256.
[
|
| [33] |
|
| [34] |
|
/
| 〈 |
|
〉 |



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}