
黄河三角洲柽柳光合作用及树干液流对潜水埋深的响应
作者简介:任冉冉(1993- ),女,山东泰安人,硕士,研究方向为生态治理与修复。E-mail: xiaoran2822@163.com
收稿日期: 2019-05-27
要求修回日期: 2019-09-04
网络出版日期: 2019-12-28
基金资助
国家自然科学基金项目(31770761)
山东省重点研发计划项目(2017GSF17104,2017CXGC0316)
山东省农业科技资金项目(2019LY006)
泰山学者工程专项
Response characteristics of photosynthesis and sap flow parameters in Tamarix chinensis leaves to depth of groundwater table in the Yellow River Delta
Received date: 2019-05-27
Request revised date: 2019-09-04
Online published: 2019-12-28
Copyright
为揭示黄河三角洲柽柳(Tamarix chinensis)叶片光合作用及耗水特征对潜水埋深的响应规律,明确维持柽柳较高光合效率及适宜生长的潜水埋深,以三年生柽柳苗木为试验材料,模拟设置淡水条件下0 m、0.3 m、0.6 m、0.9 m、1.2 m、1.5 m、1.8 m共7个潜水埋深,测定分析柽柳叶片气体交换参数的光响应过程和树干液流日动态。结果表明:不同潜水埋深可显著改变土壤水分条件,从而影响柽柳的光合作用和耗水性能,柽柳叶片净光合速率、光合光响应参数、水分利用效率以及树干液流速率具有明显的水位响应性。在浅水位(≤0.3 m)导致渍水胁迫和深水位(1.8 m)导致干旱胁迫时,柽柳幼苗光合作用受到较大抑制。柽柳呈现出耐干旱不耐水湿的光合水分适应性,深水位(≥0.6 m)柽柳的光合能力显著高于浅水位(≤0.3 m),在潜水埋深0.9~1.5 m之间柽柳光合能力较强,1.2 m是柽柳生长最适宜的潜水埋深。
任冉冉 , 夏江宝 , 张淑勇 , 赵自国 , 赵西梅 . 黄河三角洲柽柳光合作用及树干液流对潜水埋深的响应[J]. 自然资源学报, 2019 , 34(12) : 2615 -2628 . DOI: 10.31497/zrzyxb.20191211
In order to reveal the response of photosynthesis and water consumption characteristics in Tamarix chinensis leaves to the depth of groundwater table, and define the depth of groundwater table to maintain T. chinensis higher photosynthetic efficiency and suitable growth. By taking three-year-old T. chinensis seedlings as experimental materials, a total of 7 submersible depths of groundwater table (0 m, 0.3 m, 0.6 m, 0.9 m, 1.2 m, 1.5 m and 1.8 m) was designed, and the light response processes of gas exchange parameters and the daily dynamics of sap flow in T. chinensis leaves to different depths of groundwater table were analyzed. The results showed that different depths of groundwater table could significantly change the soil moisture condition, and affect the photosynthetic process and water consumption in T. chinensis leaves. The net photosynthetic rate (Pn), photosynthetic response parameters, water use efficiency (WUE) and sap flow rate in T. chinensis leaves had obvious response to the depth of groundwater table. (1) With the depth of groundwater table rising, the Pn, WUE, transpiration rate, and sap flow rate increased first and then decreased, which showed the maximum value at the depth of groundwater table of 1.2 m with relative soil water content of 40.51%. The stomatal conductance, stomatal limit value increased first and then decreased, while the intercellular CO2 concentration was the opposite; From 1.2 m to 1.8 m depth of groundwater table or from 1.2 m to 0 m depth of groundwater table, the photosynthetic decrease was mainly due to the stomatal limitation. (2) With the depth of groundwater table rising, the maximum net photosynthetic rate, light saturation point, and apparent quantum efficiency in T. chinensis leaves increased first and then decreased, reaching the maximum value (21.15 μmol·m-2·s-1, 1513.4 μmol·m-2·s-1, and 0.06 μmol·mol-1, respectively) at the 1.2 m depth of groundwater table, while the light compensation point was the opposite. (3) The 1.2 m depth of groundwater table was the turning point of photosynthesis from stomatal limitation to non-stomatal restriction. The stomatal regulation made T. chinensis maintain high photosynthetic characteristics under drought stress. The conclusion was that the T. chinensis showed photosynthetic water adaptability to drought-tolerant water and moisture, and the 1.2 m depth of groundwater table under freshwater conditions was suitable for the growth of T. chinensis.
Fig. 1 Schematic diagram and photo of soil columns with Tamarix chinensis图1 栽植柽柳的土柱模拟示意图和实景图 |
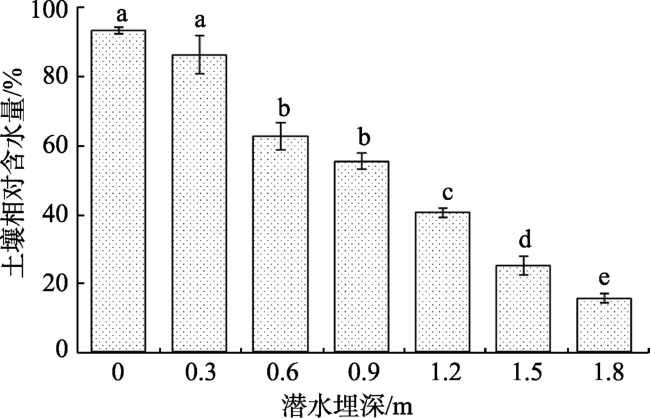
Fig. 2 Changes of soil relative water contents at different groundwater levels图2 不同潜水埋深下土壤相对含水量变化 |
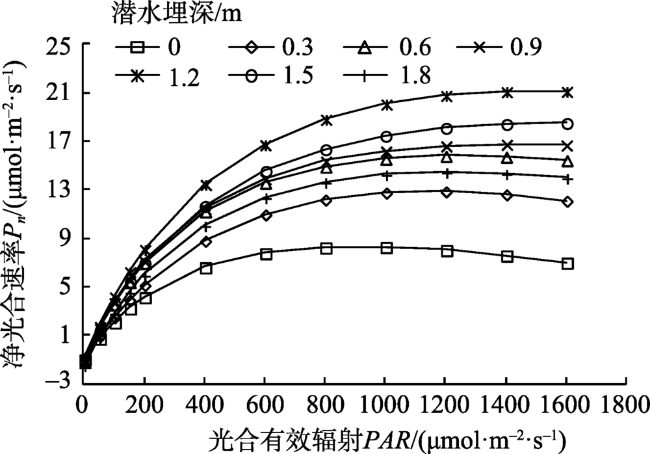
Fig. 3 Light response curves of net photosynthetic rate (Pn)of Tamarix chinensis at different groundwater levels图3 不同潜水埋深下柽柳净光合速率的光响应曲线 |
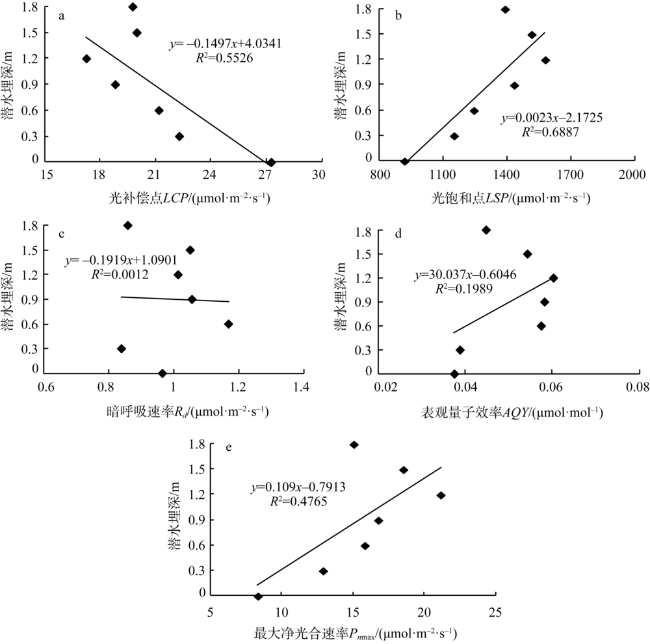
Fig. 4 The response of photosynthetic light response parameters of Tamarix chinensis to groundwater level图4 柽柳光合光响应参数对潜水埋深的响应 |
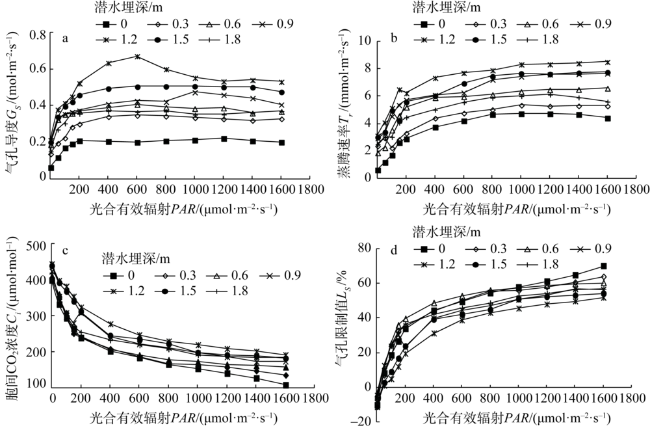
Fig. 5 Light response of stomatal conductance, transpiration rate, intercellular CO2 concentration, and stomatal limit values of Tamarix chinensis at different groundwater levels图5 不同潜水埋深下柽柳气孔导度、蒸腾速率、胞间CO2浓度和气孔限制值的光响应 |
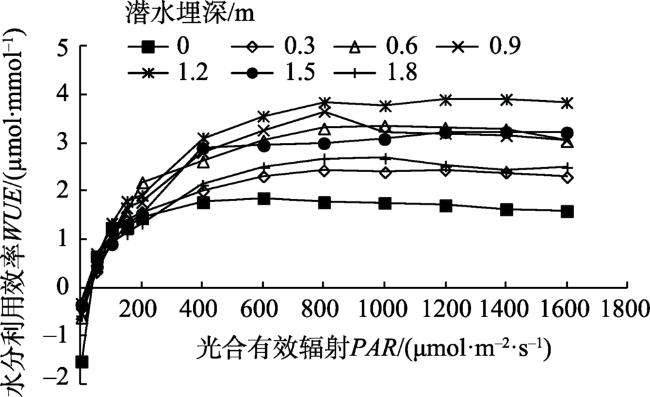
Fig. 6 Light response of water use efficiency (WUE) of Tamarix chinensis at different groundwater levels图6 不同潜水埋深下柽柳水分利用效率的光响应 |
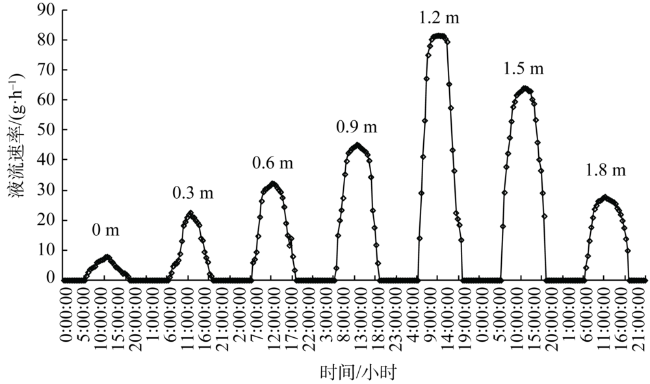
Fig. 7 Daily dynamics of sap flow velocity in Tamarix chinensis at different groundwater levels图7 不同潜水埋深下柽柳树干液流速率的日动态变化 |
| [1] |
[
|
| [2] |
[
|
| [3] |
[
|
| [4] |
|
| [5] |
[
|
| [6] |
[
|
| [7] |
[
|
| [8] |
[
|
| [9] |
[
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
[
|
| [14] |
[
|
| [15] |
[
|
| [16] |
[
|
| [17] |
[
|
| [18] |
[
|
| [19] |
[
|
| [20] |
[
|
| [21] |
[
|
| [22] |
[
|
| [23] |
[
|
| [24] |
|
| [25] |
[
|
| [26] |
|
| [27] |
[
|
| [28] |
[
|
| [29] |
[
|
| [30] |
[
|
| [31] |
[
|
| [32] |
[
|
| [33] |
[
|
| [34] |
[
|
| [35] |
[
|
| [36] |
[
|
| [37] |
[
|
| [38] |
[
|
| [39] |
[
|
| [40] |
|
| [41] |
[
|
| [42] |
[
|
| [43] |
[
|
| [44] |
[
|
| [45] |
|
| [46] |
[
|
| [47] |
[
|
| [48] |
[
|
| [49] |
[
|
/
| 〈 |
|
〉 |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}