{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
山西历山山地草甸生物多样性、优势种和土壤因素对群落生产力的影响
[赵峰侠1  , 张瑞
, 张瑞1 , 徐明2, 3 , 郑云普4 ]
, 张瑞, 郑云普|
|


作者简介:赵峰侠(1980- ),女,陕西西安人,博士,讲师,硕士生导师,研究方向为生物多样性、植物功能性状与生态系统功能的关系。E-mail: zhaofx@sxnu.edu.cn
自然生态系统生产力同生物多样性、优势种功能性状和土壤性质等多因素紧密关联,因此,受到多变量过程的综合调控。利用结构方程模型研究了山西历山山地草甸物种多样性、功能多样性、优势种功能性状和土壤等多变量因素对群落生产力的影响。结果表明:除了物种多样性外( γ=0.10, P=0.69),优势种功能性状( γ=0.61, P=0.00)、功能多样性( γ=0.47, P=0.04)和土壤因素( γ=0.34, P=0.02)直接影响群落生产力,并且土壤因素还通过改变优势种功能性状和功能多样性的方式间接影响群落生产力(间接影响系数分别为0.35、0.27)。优势种功能性状是预测群落生产力变化的最佳变量,其次是功能多样性和土壤因素。因此,理解自然生态系统生产力变化应在引入多元方法的前提下,重点考虑优势种功能性状因素。
The productivity of natural ecosystem is closely related to many factors, such as biodiversity, functional traits of dominant species and soil property, thus is controlled by multi-variable process. In this paper, the effects of species diversity, functional diversity, functional traits of dominant species and soil factors on community productivity of Lishan Mountain meadow in Shanxi province were studied by using structural equation model. The results showed that species diversity had no effect on productivity ( γ=0.10, P=0.69), but the functional traits of dominant species ( γ=0.61, P=0.00), functional diversity ( γ=0.47, P=0.04) and soil factors ( γ=0.34, P=0.02) had a direct impact on productivity, and soil factors had an indirect effect on productivity by the functional traits of the dominant species and functional diversity, with the indirect effect coefficients being 0.35 and 0.27, respectively. The functional traits of dominant species were the best predictors of productivity, followed by functional diversity and soil factors. Therefore, understanding the change of natural ecosystem productivity should be based on the introduction of multiple methods, with emphasis on the functional traits of dominant species.
生物多样性— 生产力关系是理解全球生物多样性丧失及其对生态系统功能影响的基础, 两者关系是当前生态学研究领域的热点问题之一[1]。在过去20多年里, 以Tilman为代表的数百位来自全球各地的生态学家就陆地、海洋和水生生态系统中生物多样性与生态系统生产力关系开展大量的控制实验研究, 证实生物多样性可提高生态系统生产力[2]。原因是高生物多样性使物种之间资源利用互补, 降低竞争作用, 从而增强生态系统生产力, 即资源补偿假说(Resource complementarity hypothesis)[3, 4]。以往相关研究结果表明, 基于性状的功能多样性能更好地揭示生物多样性对生态系统功能的影响[5, 6, 7], 这主要是因为功能性状对环境变化响应比物种更快[8], 其多样性比物种多样性与生产力关系更加紧密[9], 更能解释生态系统功能变化。
尽管以往研究极大地推动了人们对生物多样性— 生产力关系的理解, 但对于其研究结果及机制的解释仍存在相当多的争论。早在1998年, Grime[10]提出了质量比例假说(Mass ratio hypothesis), 认为生态系统生产力在很大程度上是由生态系统中优势种决定的, 尤其是受优势种功能性状的控制, 而对生态系统中非优势种的多样性相对不敏感, 目前该假说在全球范围内得到广泛证实[11, 12, 13, 14]。另外, 生物多样性对资源捕获的功能及其对生物量增强效应的证据主要基于控制实验即人为控制生物多样性梯度的结果[15], 因此, 该结果是否在自然生态系统成立仍存在很大争论[16, 17]。因为自然生态系统的生物多样性和生产力均受土壤养分、土壤水分、地形、气候变化和放牧干扰等多个环境因素的影响[18, 19], 可能它们对生态系统生产力的影响更为重要[20, 21, 22]。例如:在自然森林生态系统中发现生物多样性和优势种对地上生物量的作用是微弱的, 而土壤因子是影响地上生物量的重要因素[23], 主要是因为在高的土壤养分条件下, 植物生长较快, 导致地上生物量更高, 即土壤因素假说[24]。
对于自然生态系统而言, 生产力和生物多样性是由一个复杂的、多变量过程共同控制的[25, 26], 且这些过程可能同时发生[27]。因此, 不能过多关注生产力和某个驱动因素的二元模式[27, 28], 需要多变量方法来综合研究这些因素对自然生态系统生产力的影响。结构方程模型是一种用于测试一系列相关变量和独立变量直接和间接因果关系的多变量统计方法, 具有数学严谨性、描述变量之间复杂关系灵活性和直观性的优势[29]。近年来其逐渐被用于测试生物多样性、土壤、气候等多因素对森林、草地生态系统生产力的影响[22, 30], 以及用于其他方面的研究[31, 32]。基于此, 本文通过结构方程模型研究山西历山山地草甸生物多样性、优势种和土壤因素对群落生产力的影响, 旨在回答以下几个问题, 在自然生态系统中:(1)与土壤因素相比, 生物多样性和优势种功能性状是否是影响群落生产力直接的、主要的因素以及它们的影响程度; (2)去除土壤因素, 比较生物多样性和优势种功能性状对群落生产力的影响程度。研究结果将有利于深入理解生产力机制, 明确自然生态系统中生物多样性和优势种的角色, 以期为当前生物多样性丧失、生态系统功能变化的预测提供理论参考。另外, 相关结果还将为生态学中放弃简单二元模式研究、采用多元机制研究思路提供数据支撑。
历山国家级自然保护区位于山西省中条山东段, 地处沁水、翼城、阳城和垣曲四县交界, 是山西省面积最大、物种资源最丰富的自然保护区, 也是华北地区植被保存较为完整的地区之一。该区域气候属暖温带大陆性季风气候, 夏季炎热多雨, 冬季寒冷干燥, 年均温13.3 ℃, 年均降水量667.6 mm。野生种子植物达111科499属1246种。主要植被类型为暖温带落叶阔叶林, 植被垂直带自下而上主要有侧柏林、松栎林、栓皮栎林、杨桦林、山地草甸。本项目研究区位于历山自然保护区的主峰舜王坪, 植被类型为华北典型的亚高山草甸, 海拔2358 m, 其地理坐标为35° 23′ 45″~35° 25′ 25″N、111° 57′ 00″~111° 58′ 30″E, 南北长约3.2 km, 东西宽约1 km, 面积约310 hm2。土壤类型为亚高山草甸土。草甸组成以草本植物为主, 灌木较少。2007年之前由于过度放牧以及旅游等不合理开发利用导致该区域生物多样性减少和水土流失严重等生态环境问题。2007年历山国家级自然保护区通过禁牧围栏、移植草皮及播种草籽等生态恢复措施, 使该区域植被覆盖率提高到目前的80%以上, 对当地生态环境维持起着重要作用。
2017年8月中旬在植物生物量最大时调查群落。根据山体走向, 从南到北设置不同海拔梯度(2220~2320 m, 间隔为10 m左右)、不同坡向(东南、西北、东北)、不同坡度(0~15° 和30° 以上)和不同群落组成的样地12个(表1), 样地大小为50 m× 50 m, 每个样地随机布设1 m× 1 m样方3~5个, 共计44个样方。观测每个样方物种丰富度、种类、多度、盖度、频度和高度。借鉴Grime[10]对优势种划分方法将群落中相对生物量大于10%的物种确定为优势种。齐地面刈割样方的地上生物量, 去除凋落物和往年生长积累的生物量, 分种装入信封袋带回实验室, 于实验室65 ℃烘箱中烘干48 h, 称量每个物种干重用于物种相对生物量估算, 总干重用于群落生产力估算。采用5 cm直径土钻沿每个样方对角线(两个角和中心位置)采集0~20 cm土壤样品进行土壤水解氮、土壤有机碳和土壤水分含量的测定。土壤含水量采用烘干称重法, 土壤水解氮采用硼酸— 氢氧化钠碱解土壤— 扩散吸收法, 土壤有机碳采用重铬酸钾氧化外加热法。
| 表1 研究区群落组成及地形概况 Table 1 Plant community composition and topographic characteristics of the study area |
选择与群落生产力密切相关的6个地上功能性状:株高、叶经济谱(包括比叶面积、叶面积、叶干物质含量)、单株生物量、分枝数。功能性状测定参考Perez-Harguindeguy等[33]的全球植物功能性状标准化测量新手册的方法。每个样方随机选取长势良好的10株进行功能性状测定, 不足10株的物种, 测量实际株数。株高采用卷尺测量植株最大株高。每株在相同或邻近位置按不同方位成对剪下完好无损叶片(去叶柄), 放入塑封袋中, 贮藏于冰盒内带回室内。在室内, 立即用电子天平测量叶片的鲜重, 用Canon CanoScan LiDE 120扫描仪扫描叶面图像, 记录叶片数量, 随后在实验室65 ℃烘干48 h称重。用图像处理软件(Image Pro Plus)计算叶面积, 根据叶面积与其干重比值计算比叶面积, 根据叶面积与叶片数之比计算平均叶面积大小, 利用比叶面积测定过程中烘干后叶干重与叶鲜重之比计算叶干物质含量。将完整植株放入信封袋中, 在实验室65 ℃烘干48 h称重, 根据烘干重与株数比值计算单株生物量。另外, 对每一株分枝进行计数。
(1)生物多样性的计算
① 物种多样性:选用了三种常用的物种多样性指数, 即物种丰富度(R)、香农— 威纳指数(H')、Pielou 均匀度指数(J)[34], 具体计算公式如下:
式中:S为样方物种数; Pi为第i个物种个体数占群落全部个体数比例。
② 功能多样性:采用Rao系数(Rao's quadratic diversity, FDQ)计算株高、分枝数和单株生物量功能多样性[35]。具体计算公式如下:
式中:dij代表欧氏距离, 表示物种i、j在一组性状空间中的种间性状差异; Pi和Pj分别代表第i、j个物种的相对生物量。
(2)优势种功能性状值的计算
采用群落加权平均值(Community-level weight means, CWM)计算优势种株高、比叶面积、叶面积、叶干物质含量、分枝数和单株生物量的功能性状值[11]。具体计算公式如下:
式中:Pi为第i个物种的相对生物量; traiti为物种i性状值。
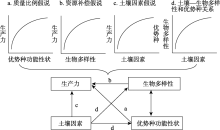
结构方程模型是基于变量的协方差矩阵来分析变量之间因果关系的一种多元统计方法, 属于验证性模型[36]。为了推断变量间因果关系, 需要先验理论知识来提出预设模型。本文根据资源补偿假说、质量比例假说和土壤因素假说, 建立预设模型(图1)。假设在自然系统中, 物种多样性、功能多样性、优势种功能性状以及土壤因素对生产力产生正效应, 并且土壤因素对优势种功能性状、物种多样性与功能多样性产生正效应。分别模拟物种多样性、功能多样性、优势种功能性状对生产力的影响, 以及它们与土壤因素相比, 对生产力的影响程度。在Amos 21.0统计软件中使用最大似然估计进行模型拟合。路径相对重要性用标准系数γ 进行比较。使用卡方值/P值(Normed chi-square, NC)和CFI值(Comparative fit index)评估模型的整体拟合效果, 判断标准为NC值在1~3之间为理想值, 小于5可接受[37]; CFI值大于0.9以上为理想[38], 统计学显著性水平标准为0.05。统计分析前, 对数据进行对数转换。整个数据处理在Excel 2010和SPSS Version 21.0软件上完成。
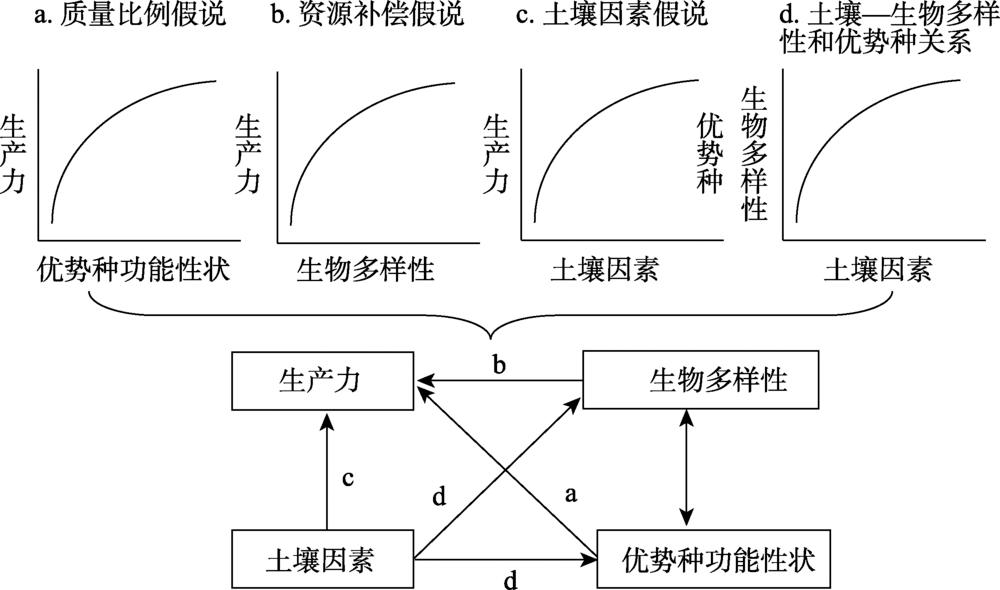 | 图1 结构方程模型的基本假设模型 注:单箭头表示两个变量的因果关系, 双箭头表示两个变量的相关关系。Fig. 1 The basic hypothesis of structural equation model |
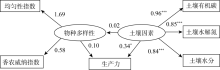
为了使模型整体拟合效果达到要求, 去除路径系数低的物种多样性与物种丰富度的路径, 得到物种多样性、土壤因素与生产力因果关系的结构方程模型NC=2.98、CFI=0.94, 表明该模型整体拟合效果很好。如图2所示, 随着物种多样性的增加, 生产力变化不明显(标准系数γ =0.10, P=0.69); 而随着土壤因素增加, 生产力呈明显增加趋势(标准系数γ =0.34, P=0.02)。同时, 随着土壤因素的增加, 物种多样性增加不显著(标准系数γ =0.02, P=0.60)。以上结果说明物种多样性对生产力的影响低于土壤因素, 并且土壤因素未对物种多样性产生影响。
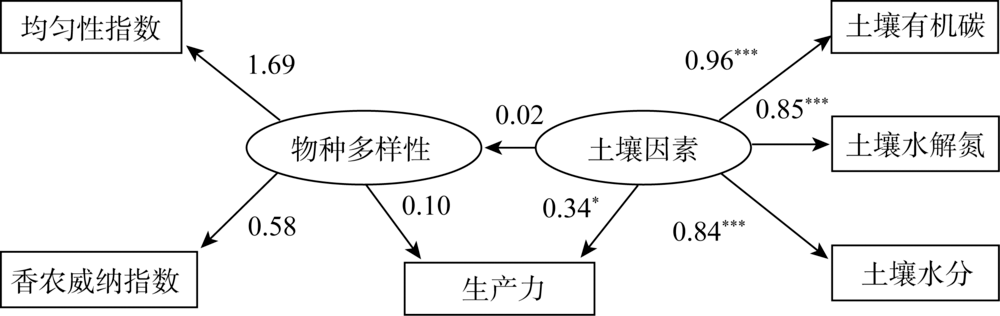 | 图2 物种多样性、土壤因素与生产力的结构方程模型 注:* P< 0.05、* * P< 0.01、* * * P< 0.001, 椭圆形中为潜变量, 矩形中为观察变量, 下同。Fig. 2 The relationship between species diversity, soil factors and productivity in structural equation model |
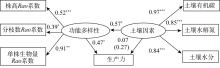
功能多样性、土壤因素与生产力因果关系的结构方程模型NC=2.23、CFI=0.90, 表明该模型整体拟合效果可接受。如图3所示, 随着功能多样性增加, 生产力也随之增加(标准系数γ =0.47, P=0.04); 而土壤因素对生产力直接影响不明显(标准系数γ =0.07, P=0.73), 但土壤因素对生产力具有间接影响(标准间接影响系数=0.27)。同时, 随着土壤因素的增加, 功能多样性增加很明显(标准系数γ =0.57, P=0.02)。以上结果表明功能多样性对生产力的影响高于土壤因素, 而土壤因素通过对功能多样性的影响而间接影响生产力。
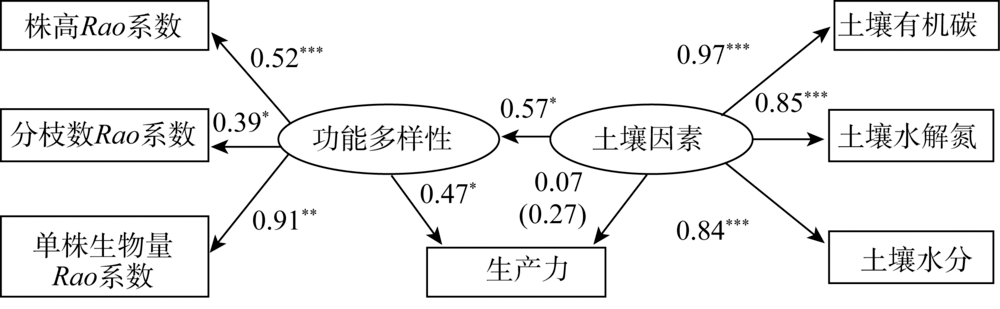 | 图3 功能多样性、土壤因素与生产力的结构方程模型 注:括号内数据代表间接影响系数, 下同。Fig. 3 The relationship between functional diversity, soil factors and productivity in structural equation model |
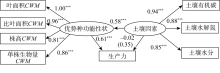
优势种功能性状、土壤因素与生产力因果关系的结构方程模型NC=3.30、CFI=0.90, 表明该模型整体拟合效果可接受。如图4所示, 随着优势种功能性状值的增加, 生产力呈上升趋势(标准系数γ =0.61, P=0.00); 而土壤因素对生产力无直接影响(标准系数γ =-0.02, P=0.89), 但土壤因素对生产力具有间接影响(标准间接影响系数=0.35)。同时, 随着土壤因素的增加, 优势种功能性状值增加显著(标准系数γ =0.58, P=0.00), 说明优势种功能性状对生产力的影响高于土壤因素, 土壤因素通过对优势种功能性状的影响而对生产力产生间接影响。
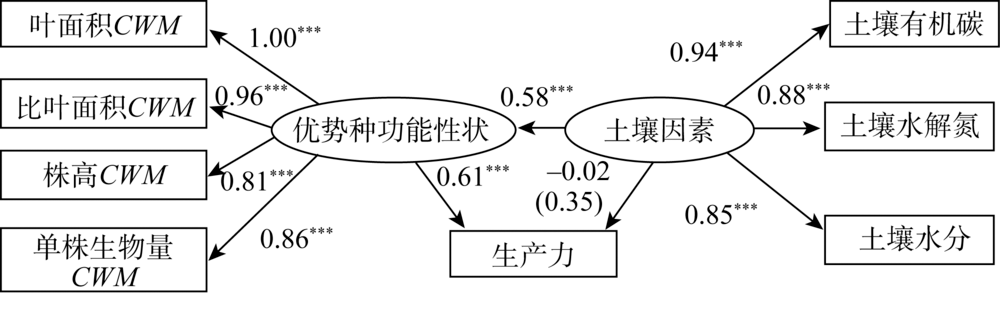 | 图4 优势种功能性状、土壤因素与生产力的结构方程模型Fig. 4 The relationship between the functional traits of dominant species, soil factors and productivity in structural equation model |
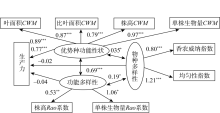
为了使模型整体拟合效果达到要求, 去除部分路径系数低的路径, 包括物种多样性与物种丰富度路径、优势种功能性状值与分枝数、叶干物质含量CWM指数路径。同时, 使用修正指标对模型进行修正, 将观察变量单株生物量Rao系数和株高CWM的误差参数由固定参数改为自由参数, 观察变量叶面积CWM和比叶面积CWM的误差参数由固定参数改为自由参数, 假设两者之间具有共变关系, 得到结构方程模型的NC=3.73、CFI=0.90, 表明该模型整体拟合效果可接受。如图5所示, 随着优势种功能性状值的增加, 生产力呈上升趋势(标准系数γ =0.77, P=0.00); 而物种多样性对生产力无显著影响(标准系数γ =-0.02, P=0.72)以及功能多样性对生产力无显著影响(标准系数γ =-0.04, P=0.62), 并且它们对生产力也无间接影响。但功能多样性与优势种功能性状值具显著正相关关系(标准系数γ =0.69, P=0.00), 以及物种多样性与优势种功能性状值也呈正相关关系(标准系数γ =0.35, P=0.01); 同时, 物种多样性与功能多样性呈正相关关系(标准系数γ =0.19, P=0.04)。说明优势种功能性状对生产力的影响高于生物多样性, 并且优势种与生物多样性相互作用, 物种多样性对功能多样性产生一定影响。
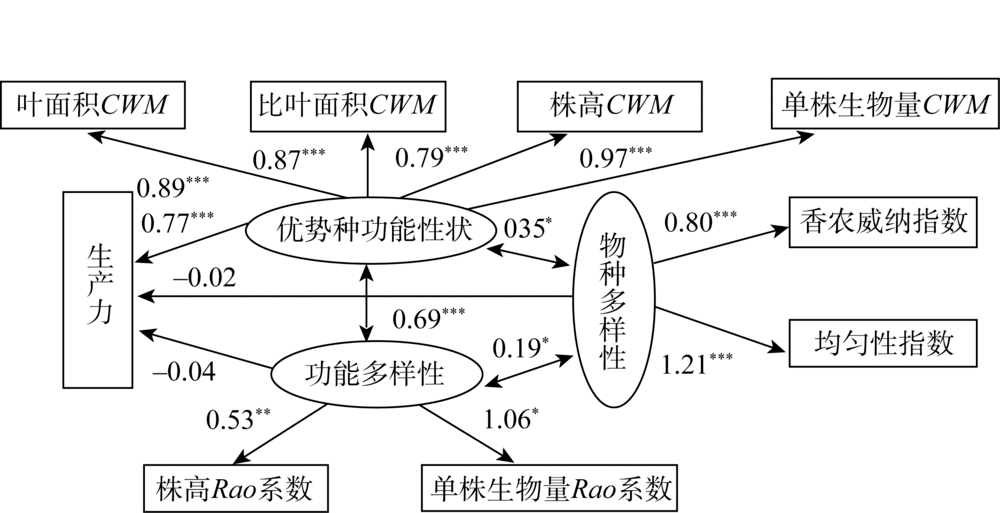 | 图5 优势种功能性状、物种多样性、功能多样性与生产力的结构方程模型Fig. 5 The relationship between the functional traits of dominant species, species diversity, functional diversity and productivity in structural equation model |
本研究发现自然山地草甸群落生产力均受优势种功能性状、功能多样性和土壤因素的影响, 证实前人提出的质量比例假说、资源补偿假说和土壤因素假说。但在对山地草甸群落生产力的贡献上, 质量比例假说占优势, 与其相关的变量对生产力来说是最好的预测者。与物种多样性和土壤因素相比, 功能多样性对群落生产力影响较大, 证实基于性状的功能多样性能更好地揭示生物多样性对生态系统生产力的影响。与物种多样性相比, 土壤因素对群落生产力影响较大, 并且土壤因素通过对优势种功能性状、功能多样性的影响对生产力产生间接影响。而物种多样性对群落生产力的影响不显著。以上结果表明, 预测全球生物多样性丧失及其对生态系统功能的影响应考虑物种功能性状, 尤其是优势种。虽然本研究在结构方程模型分析上具有一些局限性, 但证实引入多元统计方法将能更好地理解自然生态系统生产力的驱动机制。
3.2.1 生物因素与土壤因素对生产力的影响
与土壤因素相比, 物种多样性对群落生产力的影响不明显。以往相关的研究结果显示物种多样性(包括物种丰富度、物种均匀性)较之土壤养分来说对生产力的影响较弱[25, 28, 39], 与本研究结果一致。而物种多样性未表现出影响可能来自于物种间功能冗余或生态位重叠[4, 30, 40]。同时, 本研究显示土壤因素对物种多样性无显著影响, 表明物种多样性对土壤性质变化不敏感。这与以前控制实验下研究结果不一致, 如增加氮素会导致物种多样性下降[41, 42], 但与一些自然生态系统研究结果一致[28], 表明自然草地群落的物种多样性可能受气候变化[43, 44]、放牧干扰[45, 46]等其他环境因素的影响, 或者受土壤资源供应和养分相互作用等生态过程的影响[28]。
与土壤因素相比, 功能多样性对生产力具有直接影响, 这证实资源补偿假说即高生物多样性导致高资源利用效率, 从而导致高生产力[3, 25]。但与功能多样性相比(图3), 土壤因素未对生产力产生直接的影响, 而是通过对功能多样性的作用间接影响着生产力。土壤因素的这种间接效应在科尔沁草原等研究中也被证实[22, 47], 表明随着土壤因素增加, 功能多样性效应提高[48], 可能是由于土壤养分高的生境中物种间竞争排斥作用较弱[49], 从而使得物种间补偿作用对生产力影响增强。同时, 土壤因素也是驱使功能多样性变化的重要因素之一, 可以在群落水平上对植物的功能性状进行调节[50]。
与土壤因素相比, 优势种功能性状对生产力影响较强, 这证实Grime[36]提出的质量比例假说和以前的研究结果[22, 51]。大多数群落是由一些高产的物种占优势, 这些优势种具有生长速率快或资源利用效率高(如:高叶面积和株高)的特征, 因此, 单个物种就能导致生产力增加[52]。上述现象已在水生[53]和陆地[54]生态系统被证实。而与优势种功能性状相比, 土壤因素未对生产力产生直接的影响, 但其通过对优势种功能性状的影响而对生产力产生间接影响。表明随着土壤因素增加, 进一步促进优势种的生长速率或资源利用效率, 从而增加优势种功能性状的效应。另外, Hillebrand等[55]通过Meta-analysis显示在陆地和水生生态系统中, 增加养分供应会降低群落均匀性。换句话说, 就是土壤养分的增加将使优势种出现几率更高, 从而引起生产力受优势种影响较为明显。同时, 当资源供应不平衡时, 也会促进优势种增加[28], 而本研究中土壤资源不平衡性未测, 因此, 需进一步证实优势种的优势是否是由于资源不平衡引起的, 从而导致生产力的提高。
以上结果和分析表明对土壤因素等环境变化越敏感的生物因素, 可能对生态系统功能的影响更强。由于物种对环境的响应远远低于功能性状, 因此, 从基于性状的功能多样性角度研究生态系统生产力或生态系统功能调控机制更有价值, 尤其是在局部生境。
3.2.2 生物因素对生产力的影响
当所有的生物因素都被包含在结构方程模型时, 优势种功能性状对生产力影响仍是显著的(γ =0.77), 而功能多样性和物种多样性对生产力影响不显著。这与质量比例假说在自然生态系统中扮演关键作用的观点一致[56, 57, 58]。大多数评价功能性状对森林和草地地上生物量或生产力影响的研究更多支持质量比例假说, 而不是资源补偿假说[18, 22, 51, 58]。表明与物种多样性、功能多样性和土壤因素相比, 质量比例假说相关的变量能更好地预测生态系统生产力。但Paquette等[30]结果显示在养分贫瘠或恶劣气候条件下, 资源补偿效应对生态系统过程更重要, 而在高产生境中竞争排斥往往有利用少量优势种、高产物种, 从而导致优势种效应更重要, 表明生产力对以上两种机制的依赖取决于生境条件。资源补偿假说、质量比例假说一定程度上反映群落捕获资源方式以及将这种资源转化为生物量能力。在本研究发现随着土壤养分增加, 均会增加优势种功能性状和功能多样性对生产力的影响, 同时优势种功能性状值与功能多样性呈较强正相关关系(标准系数γ =0.69), 与物种多样性相关性次之(标准系数γ =0.35)。这表明群落内两种资源捕获方式并不是排斥的, 可以相互作用。当优势种功能性状越强时, 空出生态位更容易导致优势种和非优势种之间建立补偿关系, 增加生物多样性。但在对生态系统功能的贡献上, 还是以质量比例假说占优势。物种多样性与功能多样性呈正相关关系(标准系数γ =0.19), 这也在其他研究中被证实[22, 30], 说明虽然可能存在功能冗余, 但该群落中物种多样性在决定功能多样性中仍扮演着重要的作用。
致谢:非常感谢山西师范大学地理系的唐学娟、宋淑婷、裴晓霞、刘婧、温杰、王浩理、郭凯等同学在实验数据获取过程中给予的帮助, 同时, 也非常感谢历山自然保护区工作人员给予的支持和帮助。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|