
{kind=link}
{kind=link}
三个主要树种单木生物量及其器官分配模型
[王冬至1a, 2  , 张冬燕
, 张冬燕1a, 1b , 蒋凤玲1a , 许中旗1a , 张志东1a, 2 , 黄选瑞1a, 2, * ]
, 张冬燕]
|
|


第一作者简介:王冬至(1984- ),男,河北景县人,博士,讲师,研究方向为森林资源可持续经营与管理。E-mail: wangdz@126.com
目前已有不同方法构建生物量相容性模型,但基于非线性似乎不相关回归估计法实现不同树种生物量模型相容性及各器官生物量分配模型的研究较少。因此,论文以塞罕坝华北落叶松、油松、白桦3种林分为对象,基于非线性似乎不相关回归估计法和广义多项Logit模型,建立了包含哑变量的非线性可加生物量模型及各器官生物量分配模型。结果表明:不同树种树干生物量模型确定系数均大于0.90,树枝、树叶、树根生物量模型确定系数在0.77~0.93范围内,各器官生物量均方根误差和绝对误差分别在2.68~17.19 kg/株和0.83~1.39 kg/株范围内,经过检验不同树种各器官生物量模型均能满足精度需求。不同树种广义多项Logit分配模型,通过似然比检验、比分检验和Wald检验均达到显著水平( P<0.001),各器官参数均表现为显著水平( P<0.05)。不同树种树干、树枝、树叶及树根生物量比例分别在0.76~0.87、0.07~0.11、0.02~0.07、0.04~0.07范围内。包含哑变量的非线性似乎不相关生物量模型及广义多项Logit各器官分配模型,实现了生物量模型在不同树种间的通用性,并对森林生物量器官分配格局研究提供了科学参考。
, ZHANG Dong-yan
At present, there are different methods to construct compatible models of biomass. However, there are few reports on the compatibility of models for different tree species and the allocation model of component-specific biomass based on the nonlinear seemingly unrelated regression method. In this paper, the research objects were Larix principis-rupprechtii, Pinus tabuliformis and Betula platyphylla forests in Saihanba National Forest Farm, Hebei Province, China. Based on the nonlinear seemingly unrelated method and the generalized multinomial Logit model, the nonlinear additive models of biomass with dummy variables and allocation models of biomass in organs were established. The results showed that the discriminant coefficients ( R2) of stem biomass models for different tree species were all higher than 0.90, and the root mean square errors ( RMSE) and absolute errors were in the range of 16.68-17.19 and 0.84-1.07; in the biomass models of branch, leaf and root, the discriminant coefficients ( R2) were in the range of 0.77-0.93, and the root mean square errors ( RMSE) and absolute errors were in the range of 2.58-12.18 and 0.83-1.39. After inspection, the biomass models of different tree species can meet the precision requirement. The generalized multinomial Logit allocation models of different species all reached a significant level ( P< 0.001) after Likelihood ratio test, Score test and Wald test. The parameters of each organ were significant ( P < 0.05). The proportions of biomass in trunks, branches, leaves and roots of different tree species were in the range of 0.76-0.87, 0.07-0.11, 0.02-0.07 and 0.04-0.07, respectively. The models of nonlinear seemingly unrelated regression with dummy variables and the generalized multinomial Logit model of biomass allocation in organs can be used universally among different tree species and provide scientific basis for the study of forest biomass allocation pattern.
森林生物量在研究森林生态系统结构与功能[1]、生产力和生产潜力评价[2]、物质循环和能量流动[3, 4]、碳循环和气候变化影响与适应[5]中具有重要意义。然而不同器官生物量及分配规律通常会受到自然因素(光合作用、呼吸作用、枯损与生长等)和人为因素(采伐、修枝等)的影响[4], 因此, 世界各国越来越重视对森林生物量的估计、监测和评价[6], 其中生物量建模是项重要基础工作。在树木生长过程中, 各器官生物量生长均呈现异速生长规律[7], 并在生态学和应用环境学研究中广泛应用[8, 9, 10]。不同器官生物量与分配是表征立木生长过程的重要指标[11], 对了解树木变异、预测林火、木材生产及养分循环等[12]具有重要作用, 因此对立木不同器官生物量估计及其分配的研究是必须的。
目前国内外学者已对生物量建模工作做了部分研究, 汪珍川等[13]、付甜等[14]基于幂函数模型构建了亚热带主要森林类型地上各器官生物量模型; 然而由各器官生物量之和估计总生物量时预测误差较大[14, 15]。常用度量误差法、转换因子连续函数法和加权生物量回归模型[16, 17]等方法, 实现各器官生物量模型与总生物量模型之间的相容性。此外, Parresol[18]提出的非线性似乎不相关法, 能有效解决不同器官生物量方程的可加性问题。考虑到生物量模型形式、误差结构、可加性模型构造等因素对预测精度的影响[19], 董利虎等[20, 21]基于似然分析法和刀切法确定了生物量模型误差结构, 并采用似乎不相关回归方法对方程参数和偏差校正因子进行联合估计[22], 建立了以对数转换为基础的可加性生物量方程系统。符利勇等[6]和肖生苓等[23]分别采用权函数、比例平差法、非线性似乎不相关回归法和非线性联立方程组法, 来消除方程的异方差和度量误差, 建立了兴安落叶松(Larix gmelinii)和马尾松(Pinus massoniana)总生物量与各器官的相容性生物量模型。此外, 还有学者采用混合模型和误差变量联立方程组方法[24, 25, 26], 构建了不同区域、不同起源立木生物量的通用性方程。
鉴于当前国内外有关生物量预测模型的研究主要集中在总生物量等于各器官生物量之和这一逻辑关系的探究, 常用建模方法主要有可加性生物量法、联立方程组方法、似乎不相关法、混合模型法等统计方法, 其中非线性似乎不相关法是估计各器官生物量的有效方法。然而基于非线性似乎不相关回归估计法, 在一个可加性生物量模型中, 同时估计出不同树种间各器官相容性的生物量模型参数及各器官分配模型的研究鲜为少见。因此, 本文以河北省塞罕坝机械林场华北落叶松(Larix principis-rupprechtii)人工林、油松(Pinus tabuliformis)人工林、白桦(Betula platyphylla)次生林为研究对象, 首先基于非线性似乎不相关回归估计法建立包含树种效应相容性生物量模型, 然后基于广义多项Logit模型建立不同树种各器官生物量分配模型, 为构建不同林分类型或起源的相容性生物量模型及其分配规律研究提供科学依据。
塞罕坝机械林场总场(41° 22′ ~42° 58′ N, 116° 53′ ~118° 31′ E)位于河北省最北部, 与内蒙古克什克腾旗和多伦县的部分地区接壤, 属阴山山脉与大兴安岭余脉的交接地带。林区属华北暖温带立地类型区域, 海拔1 010~1 940 m, 年均气温-1.2 ℃, 年均最高气温33.4 ℃, 年均最低气温-43.3 ℃; 年均日照时数2 548.7 h; 年均降水量约452.2 mm[27]。研究区主要乔木树种有华北落叶松、白桦、云杉(Picea asperata)、樟子松(Pinus sylvestris var. mongolica)、山杨(Populus davidiana)、油松、蒙古栎(Quercus mongolica)等, 主要灌木树种有绣线菊(Spiraea salicifolia)、沙棘(Hippophae rhamnoides)、胡枝子(Lespedeza bicolor)、山刺玫(Rosa daverica)、稠李(Prunus padus)、华北忍冬(Lonicera tatarinowii)、大叶小檗(Berberis ferdinandi-coburgii)、栓翅卫矛(Euonymus phellomanus)等, 主要草本植物有蒲公英(Herba taraxaci)、野蔷薇(Rosa multiflora)、曼陀罗(Datura stramonium)、苔草(Carex tristachya)、草地老鹳草(Geanium daharicum)、藜芦(Veratrum nigrum)、地榆(Sanguisorba officinalis)、唐松草(Thalictrum aquilegifolium)等。
分别于2013— 2015年7— 8月在河北省塞罕坝机械林场, 共设置临时样地82块(其中华北落叶松人工林29块, 油松人工林26块, 白桦次生林27块), 样地大小均为30 m× 30 m。按照传统经营方式, 对不同林分类型样地均已经营过2~4次, 其中, 华北落叶松、油松及白桦的地位指数分别在7.6~25.8、4.7~23.6、5.8~21.9 m范围内。观测样地立地因子(海拔、坡度、坡向等)并对样地内立木进行每木检尺。不同林分类型样地主要林分因子见表1。根据检尺结果在每块样地内分别选取1株平均木测定其生物量。树干生物量以1 m为一个区分段进行树干解析, 分别测定每个区分段的鲜重; 枝、叶生物量采用标准枝法, 按树冠长度将其平均分为上、中、下三层, 分别在每一层按树枝基径和长度选取标准枝, 测定标准枝鲜重及标准枝上所有叶鲜重, 用每层标准枝和枝条数量来计算枝、叶生物量; 树根生物量则采用全挖法, 根据直径大小将树根分为:小根(< 2 cm)、中根(2~5 cm)、大根(> 5 cm)3组分别称鲜重。然后在树干各区分段中间截取3~5 cm厚的圆盘, 称取鲜重并作为样品烘干, 其他各部分器官均取样1 000 g作为样品, 在85 ℃恒温下烘干至恒重, 根据器官样品鲜重和干重分别推算出样木各部分干重并汇总得到单木总干重。
| 表1 不同林分类型样地基本统计量 Table 1 Basic statistical information of sample sites of different forest types |
树木生物量异速生长模型的基本形式可描述为
式中:Wgan、Wzhi、Wye、Wgen、Wguan、Wtd和Wt分别代表单木的树干、树枝、树叶、树根、树冠、地上及总生物量(kg/株); DBH为胸径(cm); H为树高(m); a、b、c和d分别为模型参数; I1、I2、I3为表示不同树种的哑变量; ε 为误差项。
在回归分析中, 若因变量是取值范围在(0, 1)区间的百分数时, 即因变量服从Beta分布[12], 利用经典线性回归(普通最小二乘法和广义最小二乘法)进行预测研究时, 由于比例数据之间存在异方差[15], 其预测值通常在(0, 1)区间之外, 预测效果较差。利用Beta分布回归模型对因变量为比例的数据进行建模, 能够克服以上问题且所产生的均方根误差和绝对偏差均较小。在林业相关研究中, 常运用Beta分布回归估计林冠郁闭度[29]和灌木盖度[30], Poudel等[12]基于Beta分布回归建立了花旗松(Pseudotsuga menziesii)和美国黑松(Pinus contorta)不同器官生物量分配与预测模型。立木各器官生物量具有较大的相关关系[15], 假设立木总生物量为1, 各器官分配比例在(0, 1)区间范围内, 并服从Beta分布, 其密度函数为:
式中:y ~ Beta(0, 1);
多项Logit模型是在响应变量类别之间不存在序次关系时[14], 基于最大似然估计法来预测各类别发生概率[30], 多用于经济学领域概率估计, 在生态学领域应用较少。本研究基于Beta分布的广义多项Logit模型来建立单木水平各器官生物量分配比例模型, 保证器官生物量概率分布在(0, 1)区间内且各项之和为1, 不同器官生物量分配模型可用以下公式表示:
式中:pgan、pzhi、pye、pgen分别为树干、树枝、树叶、树根的生物量占总生物量比例; ai、bi、ci为模型参数; x为各器官生物量(kg/株)。
基于常见统计量, 对不同模型拟合精度和预测精度进行比较, 即确定系数(R2)、均方根误差(RMSE)、绝对误差(Bias)及赤池信息准则AIC指标, 对模型精度进行检验, 不同统计量计算公式如下:
$RMSE=\sqrt{\sum\limits^{n}_{i=1}(y_{i}-\hat{y}_{i})^{2}/(n-p)}$(9)
式中:yi、
数据处理及模型建立分别采用SPSS 21.0统计分析软件及SAS 9.2中PROC SORT、PROC LOGISTIC及PROC MODEL中似乎不相关过程完成, 文中图片均采用Origin 9.0完成。
基于包含哑变量的非线性似乎不相关方法, 建立了不同树种单木的各器官可加性生物量模型, 模型参数估计值、均方根误差、绝对误差及确定系数见表2。不同林分类型树干生物量模型拟合精度最高, 枝、叶、根生物量模型拟合精度稍低, 不同林分类型各器官生物量残差分布如图1所示。不同树种树干生物量模型的确定系数均大于0.90, 均方根误差和绝对误差分别在16.68~17.19 kg/株和0.84~1.07 kg/株的范围内; 枝、叶、根生物量模型确定系数在0.77~0.93范围内, 均方根误差和绝对误差分别在2.58~12.18 kg/株和0.83~1.39 kg/株的范围内。对预测值进行残差分析(图1), 预测结果在[-95%, 95%]置信区间内, 预测值接近于真实值, 不同林分类型各器官生物量模型均能满足精度需求。
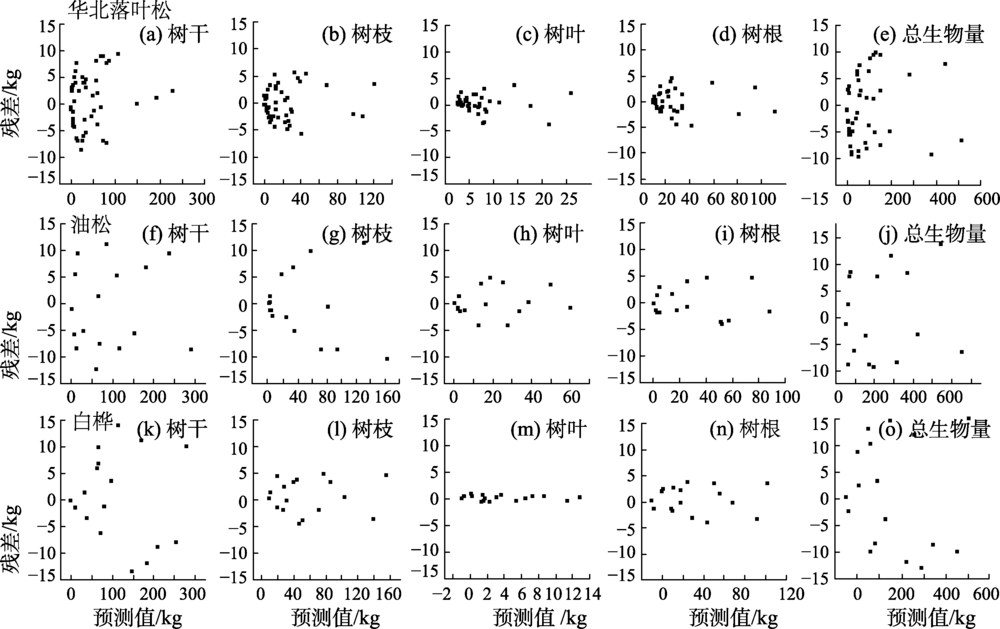 | 图1 不同林分类型各器官生物量残差分布Fig. 1 Residual distribution of biomass in various organs of different forest types |
| 表2 不同林分类型生物量参数估计及统计检验 Table 2 Parameter estimation and goodness-of-fit statistics in biomass estimation of different forest types |
树木各器官分配比例服从Beta分布, 基于广义多项Logit模型对不同树种各器官分配比例模型进行了拟合, 不同林分类型各器官生物量比例模型参数估计、参数标准误、信息准则(AIC值)如表3所示。经过似然比检验(Likelihood Ratio Test, LR)、比分检验(Score Test)和Wald检验对不同林分类型各器官比例模型进行检验, 不同林分类型各树种3种检验方法均达到显著水平(P< 0.001), 不同器官各参数均表现为显著水平(P< 0.05), 表明不同林分类型生物量分配比例模型拟合效果较好。
| 表3 不同林分类型各器官生物量比例模型参数估计与统计检验 Table 3 Parameter estimation and goodness-of-fit statistics of proportion model of organ biomass in different stand types |
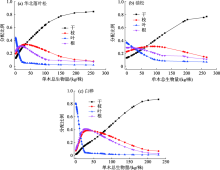
不同林分类型单木各器官生物量分配预测如图2所示, 其中华北落叶松人工林的干、枝、叶、根生物量分别占单木总生物量的0.13~0.81、0.19~0.07、0.45~0.05、0.23~0.07; 油松人工林的干、枝、叶、根生物量分别占单木总生物量的0.13~0.76、0.24~0.11、0.34~0.06、0.29~0.07; 白桦次生林干、枝、叶、根生物量分别占单木总生物量的0.02~0.87、0.08~0.07、0.81~0.02、0.09~0.04。随着单木生物量逐渐增加, 不同林分类型树干生物量比例稳定在0.76~0.87的范围内, 树枝生物量比例稳定在0.07~0.11的范围内, 树叶生物量比例稳定在0.02~0.07的范围内, 树根生物量比例稳定在0.04~0.07的范围内。
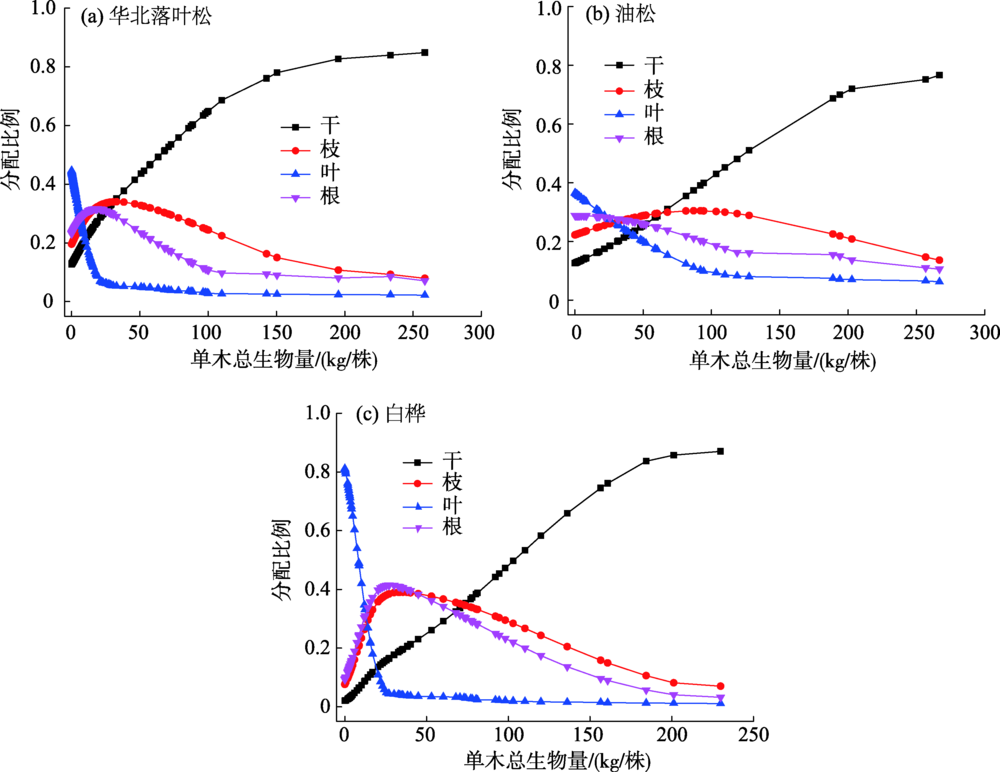 | 图2 不同林分类型各器官生物量分配规律Fig. 2 Distribution of biomass in various organs of different forest types |
在生物量预测模型研究过程中, 解决不同树种各器官生物量相容性及其生物量分配规律一直是该领域的一个难题。研究中采用包含哑变量的非线性似乎不相关回归估计法, 建立了不同树种生物量预测模型, 该方法不仅考虑了单木总生物量与各器官生物量之间的内在相关性, 而且确保各器官生物量预测值之和等于总生物量, 其模型参数估计更有效[18, 22]。将不同树种以哑变量方式加入到模型中, 同时将总生物量分为地上和地下(树根)两部分, 地上生物量分为树干和树冠两部分, 树冠生物量分为树枝和树叶两部分, 构建了包含哑变量的可加性生物量预测模型。通过分级控制建立单木总生物量模型, 能够有效消除度量误差, 解决生物量总量模型和各器官生物量模型不兼容的问题[23]。考虑不同树种的相容性生物量模型, 研究中将树种作为哑变量来反映不同树种之间差异。符利勇等[25]通过哑变量方法基于不同数量的自变量构建了相容性生物量模型, 曾伟生等[26]利用线性混合效应模型实现了不同起源树种生物量模型通用性, 而非线性混合效应模型联立方程组法则缺乏相应的算法[25]。Wang等[31]研究表明选择哑变量方法和混合模型方法均可实现模型通用性且预测结果相同。因此, 与传统方法相比, 构建包含哑变量的非线性似乎不相关生物量预测模型既能保证模型预测精度, 又能实现预测模型通用性。
不同树种各器官生物量模型中, 树干预测模型拟合效果最好, 不同树种树干模型确定系数在0.94~0.97范围内, 均方根误差在16.68~17.19 kg/株范围内, 绝对误差在0.84~1.07 kg/株范围内; 而树枝、树叶、树根生物量模型拟合精度略低, 模型确定系数在0.77~0.93范围内, 均方根误差和绝对误差分别在2.58~12.18 kg/株和0.83~1.39 kg/株的范围内, 拟合结果仍能够满足预测要求。产生误差的原因可能是由于抽样误差、样本数量偏小、树种生长过程不同、林分结构差异及生物量分配等[12, 32]影响, 导致数据变幅较大, 使部分器官产生了较大估计偏差。若想得到更精确的结果, 可能需在模型中加入立地条件、林分密度、不同发育阶段等能体现生物量差异的变量, 建立相应的指标权重体系[14], 以实现对生物量较为精确的估测。
树木各器官生物量直接反映光合产物积累量, 并影响最优化分配格局[33]。不同树种各器官分配比例不同, 树干生物量比例为0.76~0.87, 树枝生物量比例为0.07~0.11, 树叶生物量比例为0.02~0.07, 树根生物量比例为0.04~0.07, 与部分学者[7, 13, 14, 17, 33]研究结果相似。树木各器官生物量分配比例是动态变化的[33], 通常会受到诸多生物学和非生物学因素限制, 如光照、水分、养分、年龄、密度及立木大小等[4, 34]。在生长过程中, 林木会不断调节各器官生物量比例以最大化获得受限资源[35, 36]。立木对光照、水分和养分竞争能力的高低是通过各器官生物量分配实现的[37], 其中枝、叶、根生物量分配光照、水分和养分条件有显著相关性[38], 当光照成为主要限制因子时, 立木将更多生物量分配给枝和叶, 以尽可能多的获取光照; 当水分或养分成为最主要的限制因子时, 更多生物量分配给根, 以获取更多土壤水分或养分[39, 40, 41]。立木大小和密度导致树木间产生空间和资源竞争, 从而影响树木的可塑性, 进而导致各器官的生物量分配比例变化[42, 43]。在无竞争存在时, 会增加地上各器官生物量分配[17], 树干在立木组分中占有较大比重[23], 但树干生物量分配比例在不同龄组间差异却非常显著[7]。因此, 在今后研究立木各器官生物量分配过程中, 应与经营措施、立地类型、立木生长环境及不同发育阶段等结合起来并做进一步深入研究, 可能是全面理解树木异速生长机制的突破口。
致谢:感谢两名匿名审稿专家对本文提出了中肯的修改建议。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|