{kind=link}
{kind=link}
{kind=link}
生产力与生物多样性关系的研究进展
[赵峰侠1  , 徐明
, 徐明2, 3, * ]
, 徐明]
|
|


作者简介:赵峰侠(1980- ),女,陕西西安人,博士,讲师,硕士生导师,研究方向为生物多样性、植物功能性状与生态系统功能的关系。E-mail: zhaofx@sxnu.edu.cn
生产力和生物多样性是生态系统两个基本属性。然而,几十年来,人们一直在争论它们之间的相互关系及其潜在机制。论文综述过去有关两者关系的控制实验、meta分析、多点观测研究和潜在机制,发现两者关系主要表现为单峰关系、正相关关系、负相关关系、无关系、“U”型关系,并且关于它们关系的潜在机制也没有普遍共识。同时,两者关系及其潜在机制均对尺度具有很强依赖性。分析其原因,可能生产力与生物多样性的关系不是简单或直接因果关系,而是由相互关联的、多变量过程共同控制。因此,应放弃固定化二元模式,开展多元化驱动因素的生物多样性研究,加强人类对复杂生态系统过程的理解,提供更具意义的生物多样性保护和管理指导。
Productivity and biodiversity are two fundamental properties of ecosystems. However, there has been a long-running debate over their interrelationship and the mechanisms behind their relationship for decades. In this paper, the control experiments, meta-analysis and multi-site observation studies over the past decades for testing relationships between productivity and biodiversity and the mechanisms of their relationships are reviewed. It is found that the forms of productivity-biodiversity relationships mainly are: unimodal relationship, positive relationship, negative relationship, no relationship and U-shaped relationship. Besides, there is no general consensus about underlying mechanisms of their relationships. Meanwhile, both the relationships and the underlying mechanisms are strongly dependent on scales. These results illustrate that the productivity-biodiversity relationships are not a simple or direct causal relationship, but are driven by interconnected and multivariate ecological processes. Therefore, we should give up a fixation on bivariate patterns and adopt the approach of multivariate drivers to the problem of the biodiversity pattern in order to further strengthen our understanding of complex ecosystem processes and provide more meaningful guidance for conservation management.
生产力和生物多样性是生态系统两个基本属性[1], 两者关系是生态学中研究最多的模式之一[2]。研究两者之间的关系对于认识全球生物多样性的机制是至关重要的[3]。然而, 虽然有大量的文献研究两者关系, 但对生产力与生物多样性关系的模式及其机制的解释仍存在争议[4, 5]。正如Grace等[6]感叹“ 生态学上没有其他两个变量能像生物多样性与生产力这样被给予如此多讨论” 。目前关于生产力与生物多样性的关系出现两种研究思路:1)研究生产力对物种丰富度、群落组成和营养结构的影响, 简称“ 生产力-生物多样性关系(The productivity-biodiversity relationships, PBRs)” ; 2)研究物种数量、功能群数量、特殊物种或功能群的重要性对生产力的影响, 简称“ 生物多样性-生产力关系(The biodiversity-productivity relationships, BPRs)” 。这两种研究思路虽然在概念上有联系, 但实际研究中在很大程度上是独立的[7]。本文仅就这两种研究思路中的生产力-生物多样性关系的以下3个方面做一简要回顾和展望, 生物多样性在这里主要指物种多样性:1)关于两者关系开展的控制实验、meta分析和多点观测研究; 2)关于两者关系潜在机制的研究; 3)针对目前存在的问题提出开展多元化驱动因素的生物多样性研究, 以期为目前国内学者研究生产力-生物多样性关系提供参考。
生产力-物种多样性控制实验一般都是通过养分(如施肥)或者其他限制性因素调控生产力梯度, 从而测试不同尺度下生产力-物种多样性的关系及其机制。实验发现物种多样性随着生产力增加而发生降低[8, 9, 10, 11], 也有一些研究显示两者之间呈正相关关系或无关系[12, 13, 14]。但引起学者们广泛关注的是Grime[15, 16]和Al-Mufti等[17]最初提出生产力-生物多样性的单峰关系(The unimodal relationship)或称为“ 驼背” 模型(The“ humped-back” model, HBM)。该模型认为开始随着生产力提高, 物种多样性呈增加趋势, 随着生产力进一步提高, 物种多样性将减少, 物种多样性的峰值出现在生产力中间位置[15, 17, 18], 并且这个峰值是由两个相对过程引起的。来自陆地和水生生态系统植物、动物和微生物研究广泛支持“ 驼背” 模型[18, 19, 20, 21, 22, 23]。例如, 早期实验有Guo等[18]在草地和灌木林过渡区不同微生境中检测到“ 驼背” 模式。Moore等[19]考察加拿大东部15个湿地群落的物种丰富度和植物生物量之间的关系, 发现一个明显的单峰模式。Dodson等[20]发现33个淡水湖浮游植物和水生植物物种丰富度-初级生产力呈显著的单峰关系。因此, 一段时期内大多数研究者认为“ 驼背” 曲线是“ 正确的” 或者是“ 普遍存在的” [24, 25], 甚至被认为是经验或理论支持的唯一模型[26], 以致于后续对全球多样性模式的许多解释都是以“ 驼背” 模型作为基本假设的[27]。
“ 驼背” 模型并不总是明显的[28, 29, 30]。20世纪90年代开始, 基于大量meta分析结果, 学者们开始对生产力-生物多样性之间的“ 驼背” 模型普遍性产生质疑。生态学家通过大量meta分析从不同空间尺度、分类单位和生态系统类型的角度统计两者之间关系, 发现存在5种模式, 分别为正相关关系、负相关关系、单峰关系、无关系以及少量“ U” 型关系[2, 7, 29]。同时, 通过meta分析发现两者关系的变化依赖于尺度[7, 21, 29, 31, 32, 33, 34]。例如, Waide等[7]基于生态尺度(The ecological scales)发现群落内大多数实验结果显示两者之间无关系, 单峰模式和正相关关系次之; 在群落间得出同样结果; 然而, 在大陆到全球尺度(The continental-to-global scales)上, 正相关关系占优势。基于地理尺度(The geographic scales)即在局地尺度(< 20 km), 显示大多数是单峰模式和无关系; 在景观尺度(20~200 km)和区域尺度(200~4 000 km)出现的正相关和负相关关系多于单峰模式; 而大陆到全球尺度(> 4 000 km)中显示正相关关系占优势。Mittelbach等[29]在Waide等[7]基础上, 针对Waide等[7]研究中的方法和统计问题, 采用广义线性模型(The generalized linear models, GLIM)和普通最小二乘回归(The ordinary least squares regression, OLS)通过meta分析显示:基于生态尺度, 在群落内和群落间, 单峰模式占优势。基于地理尺度, 小于大陆的地理尺度上大多出现单峰关系, 其次, 是正相关关系, 而在最大空间尺度(> 4 000 km)单峰关系和正相关关系同时占优势。Gillman等[2]在Mittelbach等[29]基础上, 剔除受人为影响大、外来种、取样面积小、生产力被其他指标替代的研究, 结果发现正相关关系比其他关系都占优势, 该结果在Gillman等[35]后期研究中也被证实。这与以前的meta分析有实质不同, 以前研究大部分显示在小尺度上单峰模式占优势, 大尺度上正相关关系占优势[7, 29]。因此, Gillman等[2]认为尽管在他们的研究中有单峰关系出现, 但它的普遍性和重要性被夸大了。
其次, 通过meta分析发现生态系统类型也会影响两者之间的关系。水生生态系统比陆地生态系统更易产生单峰模式, 陆地生态系统中草地生态系统比森林生态系统易发生单峰模式[2, 7, 29, 36, 37]。还有一些研究显示生产力和物种丰富度的关系受数据统计方法的影响。例如, Mittelbach等[29]发现采用广义线性模型(GLIM)易产生非线性关系(如单峰和“ U” 型模式), 而采用普通最小二乘回归(OLS)易产生线性关系, 特别是正线性关系。还有一些研究发现使用的生产力指标不同也会影响两者之间的关系。目前衡量生产力指标不仅有净初级生产力(The net primary productivity, NPP)和现存生物量, 还有一些替代指标, 如实际蒸散量、潜在蒸散量、降雨量、温度、纬度、海拔和土壤养分状况等[29, 38]。不同生产力指标会随着尺度变化而发生变化, 当植物生产力用现存生物量测量时, 单峰模式占优势[30]。运用替代指标时, 其一般假设替代指标与生产力呈正相关性。然而, 在降雨和实际蒸散量等替代指标处于高水平时, 比如在暖湿条件下, 土壤营养物质在强风化和淋溶的作用下, 使得土壤养分和植物生产力不可避免地降低[38], 从而导致替代指标与NPP呈负相关性, 而不是开始假设的正相关性, 结果影响生产力-物种多样性的关系[39]。Š í mová 等[40]甚至发现当用NPP作为生产力指标时, 不同NPP估测方法也会影响生产力和物种丰富度关系, 如:基于MODIS NPP模型模拟的NPP与物种丰富度具有很好的正相关关系。鉴于meta分析出现的混合结果以及评价方法的不统一性, 学者们对目前两者关系使用meta分析持批判态度, 甚至认为应该结束生产力和物种多样性间的meta分析[1]。
鉴于大多数meta分析没有考虑研究方法的差异性, 如取样大小、生产力指标测量等, 同时, 来自自然生境的研究有时也会从meta分析中去除, 仅支持控制实验[41]。Grime等[42]认为这不能达到理解自然界发生现象的目的。因此, 继meta分析之后, 关于生产力-生物多样性关系涌现出大量来自自然生态系统的多点观测研究。例如, Adler等[37]通过标准化取样面积和生产力指标(采用地上净生产力), 研究5个大洲48个草地群落在局地、区域以及全球范围内生产力-物种丰富度关系, 结果发现在所有的尺度上生产力-物种丰富度之间无明显关系。例如, 采用标准回归和准泊松回归在48个位点中发现34个表现出无明显关系, 5个表现出正相关关系, 5个表现出负相关关系, 3个表现出“ U” 型关系, 1个是单峰关系, 因此, Adler等[37]得出结论:生产力对植物物种丰富度的影响很弱。该结论在学术界引起很大反响, Pan等[43]、Fridley等[44]和Pierce[45]分别对其结论进行评论。Pan等[43]对Adler等[37]的统计方法和结论提出质疑, 认为他们的方法偏向于位点数量多的植物群落, 如草原、荒漠草原和混合草草原分别仅有1个位点, 而一年生草地有6个位点。为了解决这个问题, Pan等[43]在回归分析中使用植被类型降低不同植物群落位点数差异, 利用每个位点植被类型平均生物量和平均丰富度来估测全球30个位点中12种植被类型生产力和物种丰富度的关系。结果显示在全球尺度上, 生产力-物种丰富度间呈显著正相关关系, 与Gillman等[2]研究结果一致。Pierce[45]采用上限回归(The upper boundary regression)重新分析Adler等[37]的数据, 得出所有位点内两者关系呈显著“ 驼背” 模式(R2=0.81), 在全球尺度上, 使用上限回归分析产生一个几乎完美的“ 驼背” 关系(R2=0.98)。因此, Pierce[45]和Fridley等[44]都认为Adler等[37]没有得到明显“ 驼背” 模型, 是因为他们的研究剔除人为影响位点和1个盐沼位点。除此之外, Fridley等[44]还认为Adler等[37]分析缺乏足够的高生产力位点以及生产力估算没有考虑凋落物生物量, 如果满足以上要求的话, Adler等[37]的数据可能会产生明显的“ 驼背” 模型。因此, Fraster等[46]运用全球6大洲、19个国家、30个位点的综合调查数据估算生产力-物种丰富度关系的形式和强度。结果显示:无论是从区域尺度还是全球尺度都支持“ 驼背” 模型。其次, Fraster等[46]发现Adler等[37]的研究最大活体生物量是1 535 g/m2, 而他们研究中活体生物量达到 3 374 g/m2。此外, 两个研究每个位点取样量也不同, Adler等[37]每个位点取30个样方, 而Fraster等[46]每个位点取128~894个样方。因此, Fraster等[46]认为他们研究中出现较多的“ 驼背” 模型, 可能是由于样本量大和生物量范围宽。
然而, Fraster等[46]结果也引起争议。Tredennick等[47]认为Adler等[37]和Fraster等[46]的分歧, 是由于Fraster等[46]的统计分析中存在问题。Fraster等[46]通过位点内的生产力梯度采用网格法进行样地布设和抽样, 但没有考虑“ 网格” 的随机效应。而Adler等[37]根据当地生产力梯度随机布置样地, 并且在采用广义线性混合模型(GLMM)进行数据分析时考虑样地间随机效应。基于此, Tredennick等[47]用考虑“ 网格” 随机效应的方法重新分析Fraster等[46]的数据, 结果发现28个位点仅有29%产生单峰关系。因此, Tredennick等[47]得出结论:如果考虑到“ 网格” 的随机效应, Adler等[37]和Fraster等[46]的结果应是一致的, 即生产力-生物多样性关系很弱。与以上研究结果不同的是, 朱桂丽等[48]通过西藏典型高寒草地1 700 km野外样带的调查, 发现物种丰富度与地上生物量间存在显著的幂指数关系(y=0.219 7x0.754 9, R2=0.61, P< 0.01)。基于以上混合结果, Grace等[4]认为这些都说明目前的研究过多关注二元模式(The bivariate patterns), 导致忽视对多元机制的深入了解, 而多元机制控制着这些重要的生态系统特性。因此, 二元模式可能阻碍进一步理解生态系统, 需要进一步论证这种简单模式得到的结论或解释的内在局限性。
生产力-生物多样性关系背后的机制很长时间处于争论之中[49], 不同的关系模式有不同的机制解释, 即使同一关系模式也有不同机制。本文在这里主要介绍目前受关注比较高的“ 驼背” 模式和正相关关系的机制以及其他一些近年来引起讨论的机制。
大多数研究者对于“ 驼背” 模型解释主要是基于竞争排斥假说(The competitive exclusion hypothesis)[8, 24, 26], 认为低生产力时胁迫和资源缺乏限制物种数量, 所以随着生产力增加, 物种丰富度上升, 而当生产力处于高位时, 由于竞争排斥增加, 从而降低物种丰富度[15, 16], 或者由于高生产力导致限制性资源异质性降低[30, 50]。然而, 该机制也遭到Abrams等学者质疑, 例如:Abrams[27]列举在竞争存在情况下3种机制可使两者之间产生正相关关系。基于抽样效应的自疏假说(The self-thinning hypothesis, STH)认为在生产力低的生境中, 多度和物种丰富度随生产力提高而增加, 因为密度不足以消耗所有可利用的空间; 但当植物个体随着生产力的提高而增大到一定程度时, 将会导致它们的多度和物种总数量降低[51]。因此, 自疏机制也导致生产力和生物多样性呈现一个“ 驼背” 模式, 但与竞争排斥原理相反, 自疏假说假设物种丧失是随机的。对于“ 驼背” 模型的解释还包含有干扰[52]、进化史和扩散限制[53]等其他机制。
物种能量假说(The species-energy hypothesis, SEH)预测生产力-物种丰富度是一种正相关关系, 特别是在大的空间尺度[52]。实际上该假说是对岛屿物种-面积理论的延伸, 认为NPP或植物总碳等生产力指标作为能量输入系统中, 通过限制个体密度来限制物种丰富度, 因为高的生产力会增加更多的个体数量, 而物种数量由总的个体数量来调控[52, 54], 因而导致物种多样性提高, 因此该假说也被称为更多个体假说(The more individuals hypothesis, MIH)。目前该假说已被一些研究证实[54, 55, 56]。进化速率假说(The rates of evolution hypothesis)也预测生产力和生物多样性间呈正相关关系, 该理论认为在高生产力环境里进化会快速发生, 从而导致更多的物种形成以及物种积累速率提高[57, 58, 59], 涉及到物种形成的进化时间概念[57]。目前已有一些证据支持分子进化速率与生产力和物种形成的关系[59, 60]。综合进化速度假说(The integrated evolutionary speed hypothesis)进一步延伸进化速率假说[35], 但与前者不同, 进化速率假说是把生产力作为一个关键变量, 而综合进化速度假说是将温度、水分以及生物群落面积作为关键变量来确定总NPP, 预测其与进化速度、物种形成速率、最终物种积累和生物多样性呈正相关性, 从而将生产力与遗传进化联系起来[35, 61]。与生物相关的水-能量动力学假说(The biological relativity to water-energy dynamics hypothesis)也用来解释生产力和生物多样性之间的正相关性。该假说认为生物动力学对能量/物质条件变化的相对性质, 是由于水的变化引起的, 尤其是液态水[62]。当液体水-能量动力在相当长一段时间内最大时, 物种地理分布的进化速率和变化速率将是最大的, 因此, 它在生态和进化过程中引起生产力和物种多样性之间的正相关关系[35, 63]。
近来还有一些受关注度比较高的机制解释生产力-生物多样性关系。例如, 异质性假说(The heterogeneity hypothesis, HH)认为高生产力与更广泛的环境条件有关, 在高产量的区域物种数量的增加是由于更高的物种空间周转或者更高生境专属化[64, 65], 但对两者关系的解释依赖于尺度[66]。可能在小的空间尺度, 其他过程(如竞争排斥或抽样效应)占优势, 而在大的空间尺度, 异质性对物种丰富度正效应变得更重要, 因为它积累了小尺度环境变化效应[67]。物种库假说(The species pool hypothesis, SPH)认为在给定区域内物种数量只是来自于能够忍受当地条件的物种总数, 被用于解释在低纬度暖湿地区(高产生境)高的物种多样性, 并且也有越来越多证据支持物种库假说[68, 69]。Huston[39]发现物种库的大小在生产力-物种多样性关系中会影响到最大多样性的值, 也就是影响“ 驼背” 模型峰值。因此, Huston[39]认为发生在当地空间尺度上, 潜在竞争物种的最大数量由区域内所有物种数量决定, 即当地或区域的物种库, 并依赖于时空尺度。
以上提到的假说基本上都未充分解释生产力-物种多样性的关系。竞争排斥假说、基于抽样效应的自疏假说、物种能量假说、进化速率假说等忽视物种的数量也受区域尺度的影响, 而物种库假说没有考虑小尺度的生物相互作用[53]。Š í mová 等[70]提供有力证据证实生产力-物种丰富度关系受多重机制驱使, 以及每个机制的影响随空间尺度会发生变化。然而, 目前大多数研究仅把焦点放在特定假说上, 忽略多重机制控制生产力-物种多样性关系的事实[71]。因此, 生产力-物种多样性关系的机制需通过多个尺度的多个假设同时测试才能更好地理解[72]。
综合控制实验、meta分析和多点观测的研究结果, 生产力-生物多样性关系没有统一模式, 说明生产力-生物多样性关系可能不是简单或直接因果关系, 这里面需要考虑到除生产力外, 气候、干扰、进化时间、生物相互作用、生态位、面积、空间异质性、物种库以及群落组成历史等因素对生物多样性的影响。正如Grace等[4]所说, 对生物多样性模式的影响是由多个相互关联过程控制着, 并且这些过程同时发生。虽然Adler等[37]和Fraster等[46]的研究结果存在分歧, 但他们都认为对生物多样性模式的解释需引入多元模式。其次, 通过对meta分析和多位点观测研究的分析, 认识到导致生产力-生物多样性关系依赖于空间尺度、分类单位或生态系统类型, 尤其是空间尺度, 可能不同空间尺度的生物多样性受多个驱动因素的影响, 或者以某个因素影响为主, 其他因素为辅。同时, 控制两者关系的机制随尺度发生变化以及在不同尺度上每种机制发挥作用也不同。还有一些需要关注的细节, 例如, 取样面积、取样点布设范围、生产力的测定方法等。此外, 气候变量中温度、降雨、蒸散量等一般在大空间尺度上与物种丰富度成呈正相关性[73, 74], 同时生产力也与这些变量存在共变性[40], 所以会产生生产力对生物多样性的影响可能是因为它与其他变量共变性引起的或者是它们之间共同影响的结果。因此, 在开展多元化驱动因素对生物多样性模式影响研究时, 还需建立一种新方法将生产力与这些变量对生物多样性的影响分离。
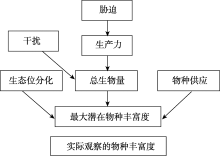
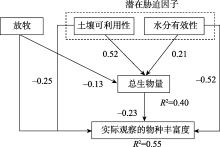
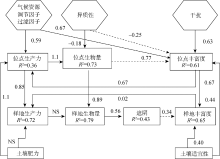
目前Grace等已开始开展多元化驱动因素的生物多样性模式研究工作, 为了敦促科学家超越“ 驼背” 模式, 放弃固定化二元模式, Grace等[6]通过因果网络(The causal networks)方式考察Grime[15, 75]提出的“ 驼背” 模型, 他们先把“ 驼背” 模型的预测翻译成因果图(图1), 这个因果图明确地表达出多个过程(包括潜在环境胁迫和干扰)影响着生物量、物种丰富度以及它们之间的关系。然后使用结构方程模型(The structural equation modelling, SEM), 结合已发表芬兰草地数据[76]评估该因果图, 结果显示(图2):代表环境胁迫因子的土壤可利用性和水分有效性以及代表干扰因子的放牧可直接影响物种丰富度。这些结果与“ 驼背” 模型基本假设不一致, 因为“ 驼背” 模型认为干扰和胁迫对物种丰富度的影响是通过总生物量来预测的。该结果还发现在模型中运用全套变量预测物种丰富度的变化(R2=0.55)要比单独使用生物量一个变量(R2=0.14)效果更好。2016年, Grace等[49]又采用结构方程模型整合多个理论到单个模型, 收集全球5大洲39个以草本为优势的位点共1 126块样方(1 m× 1 m)的数据, 评估多个过程对物种丰富度的影响(图3)。该研究不仅将多元驱动因素扩展到气候、遮阴、土壤适宜性、土壤肥力、异质性和干扰, 而且考虑到空间尺度。结果发现, 多个变量相互间的影响具尺度依赖性, 并且不是所有的因素都在所有空间和时间上起作用。其次, 发现综合模型比传统的二元模型具有更高的解释力。二元模型对观察到的物种丰富度变化的解释不超过10%, 而结构方程模型在位点间和位点内对物种丰富度变化的解释分别是61%和65%。说明考虑多元化因素的影响是重要的, 因为它更详细地表达出, 在自然界, 特定管理措施(如通过施肥增加土壤肥力)以及气候条件的变化都可能改变生产力和物种丰富度。因此, 生态学家应超越二元模式的分析, 进一步加强人类对复杂生态系统过程的理解, 提供更具意义的生物多样性保护和管理的指导[26]。
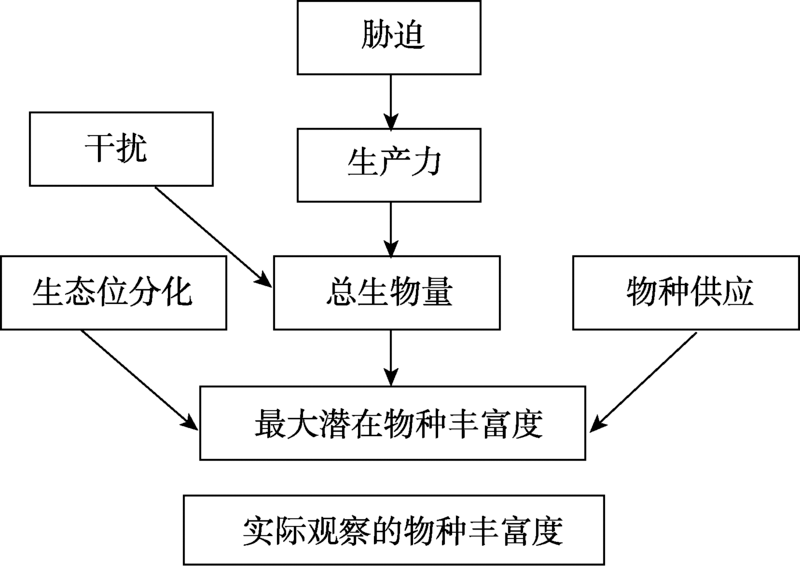 | 图1 基于Grime[15, 75]描述“ 驼背” 模型的因果图 注:来自文献[6]。Fig. 1 Causal diagram based on the “ humped-back” model described by Grime[15, 75] |
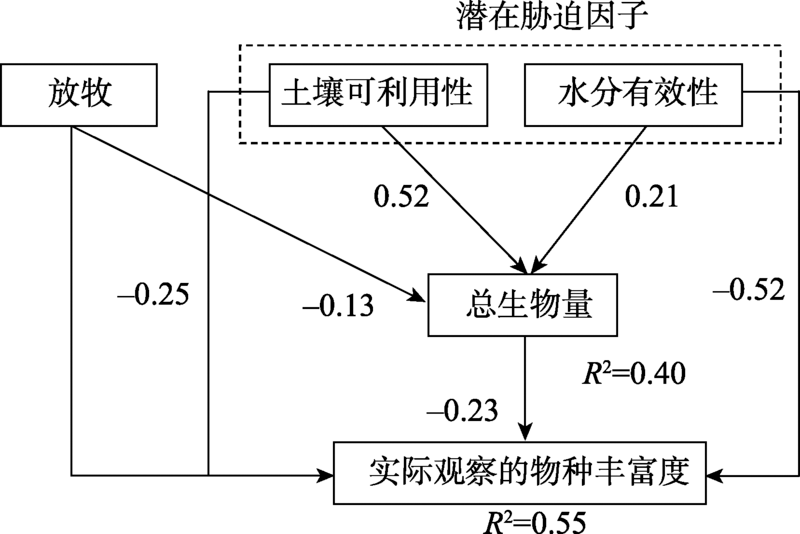 | 图2 基于Grace等[76]数据建立的结构方程模型 注:来自文献[6]。路径旁边的数字表示标准化后的影响。下同。Fig. 2 Structural equation model based on data from Graceet al[76] |
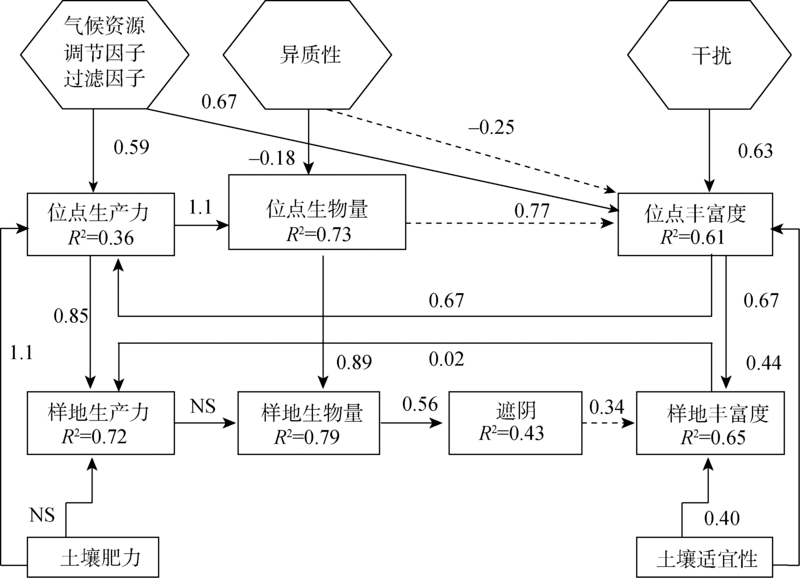 | 图3 基于全球草地数据建立的结构方程模型 注:来自文献[49]。实线箭头表示正相关, 虚线箭头表示负相关; NS表示两者关系无统计学意义。Fig. 3 Structural equation model based on data from global grassland |
与此同时, 近年来对已有生态假说的挑战, 其根本分歧在于对生态过程和理论的发展和验证[39]。过去几十年提供的生产力-物种多样性相互作用的过程可作为整个系统影响的一部分, 但需增加与此相关的其他多个驱动因素之间作用的关系, 从而使生态假说变得更为完善。此外, 关于生产力-物种多样性关系的争论也可能是由于缺乏整合思想和同时检测它们综合效应的方法。通过整合和测试它们的综合效应, 进一步促进生态理论的发展, 从而对自然系统提供系统性的理解, 提高我们预测人类改变环境因素、生产力和生物多样性的可能后果, 这样将会使生态学在未来成为一种更为精准的学科。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|