{kind=link}
{kind=link}
冬季长江口渔场游泳动物群落种类组成和多样性的年代际变化
[李建生1, 2  , 姜亚洲
, 姜亚洲1, 2 , 金艳1, 2 ]
, 姜亚洲|
|


第一作者简介:李建生(1976- ),男,副研究员,从事渔业资源评估与生态学研究。E-mail:jianshengli@sina.com
根据1991—2011年间3个代表年冬季长江口渔场的调查数据,对该海域游泳动物群落的年代际变化进行了研究。结果表明:1991—2011年,游泳动物种类数总体表现为升高的趋势,其中鱼类的增加幅度较小,甲壳类和头足类的增加幅度较大。以重量和尾数为代表的资源密度指数均表现出先下降后升高的趋势。平均体重先呈现急剧下降然后继续小幅下降的趋势。1991年优势种类数为7种,2001年为9种,2011年为7种,后2个年份和1991年相比,优势种发生了较大的更替。细点圆趾蟹( Ovalipes punctatus)和龙头鱼( Harpadon nehereus)为3个年份的共同优势种。游泳动物的站位平均种类数呈现持续升高趋势,而站位平均丰富度指数、多样度指数和均匀度指数则呈现出先升高再降低的趋势。1991和2001年、1991和2011年之间的平均相异性较高,均超过60%,而2001和2011年之间的平均相异性较低,为36.05%;引起群落差异的主要种类数呈增加的趋势,生态类群也逐渐扩大。游泳动物长度谱呈现逐渐变窄的趋势,主要是由于群落中的生物种类演替、小型鱼类数量增多和大型鱼类的群体年龄结构降低等原因造成的。
According to fishery survey data in the Yangtze estuary fishing grounds in three representative winters from 1991 to 2011, the inter-decadal variation of the nekton community in the area were studied. The results showed that the composition, the fishery resources density, the dominant species, the diversity and size spectrum of nekton species all changed significantly from 1991 to 2011 in the Yangtze estuary fishing grounds. The number of the nekton species overall showed an increasing trend, in which the number of fish species increased a little, but the number of crustaceans and cephalopods increased greatly. The density indices, including weights and individual quantity, all had increasing trend after a falling trend at the beginning, but the amplitudes of them were different. The average weight fell sharply at first and then kept dropping slightly. The number of dominant species was seven, nine and seven in 1991, 2001 and 2011 respectively, and the dominant species changed dramatically. The common dominant species in the three years were Ovalipes punctatus and Harpadon nehereus. The average number of nekton species at all stations showed a sustained increase, but the average richness, the diversity indices, and the evenness indices all firstly increased and then decreased. The average dissimilarity were higher than 60% between 1991 and 2011, 1991 and 2001. But the average dissimilarity between 2001 and 2011 was lower (36.05%). The number of main species which caused community difference was increasing, ecological groups have gradually expanded. The nekton size spectrum was gradually narrowed, mainly due to the species succession in the community, the increasing of small fish population and the decreasing of large fish population age structure etc.
长江口渔场位于长江河口外侧海域, 东至125º 00′ E, 南北分别以31º 00′ N和32º 00′ N为界。由于受长江冲淡水、苏北沿岸流、东海外海暖水、台湾暖流和黄海水团等海流水系在不同季节和年份间相互消长变化的影响, 因此该渔场水质肥沃, 饵料生物丰富, 游泳动物群落结构复杂, 具有对东黄海较为关键的海洋生态系统[1]。作为中国最大的河口, 长江口具有特殊的地理位置和环境条件, 河口生物群落在东黄海生态系统的功能中发挥着重要的作用, 尤其对海洋渔业的发展具有较大的贡献。河口水域不仅有丰富的渔业资源如凤鲚(Coilia mystus)、刀鲚(Coilia ectenes)、脊尾白虾(Exopalamoncarin cauda)等种类, 同时还是多种养殖对象苗种资源如日本鳗鲡(Anguilla japonica)、中华绒螯蟹(Eriocheir sinensis)等的分布区[2, 3]。历史上, 长江口近海水域渔业资源丰富, 其中冬汛带鱼(Trichiurus japonicus)的产量就达几十万吨, 而大黄鱼(Larimichthys crocea)的最高年产量也曾达20× 104t[1]。20世纪90年代以来, 由于东海区的捕捞努力量持续维持在高水平[4, 5]、三峡截流对长江冲淡水和生物群落的影响[6, 7]、大量发生的海水富营养化[8, 9, 10]、夏秋季东海北部海域持续爆发的大型水母[11, 12]和长江口低氧区[13, 14, 15]等因素, 对长江口游泳动物的产卵和索饵等洄游活动都产生了不同程度的影响。因此, 长江口水域已经成为国内外海洋和渔业科研工作者关注的热点水域。对于长江口及邻近水域的渔业生物资源分布、种类组成、群落结构和多样性特征已经有大量的调查和研究[16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27]。这些研究包括鱼类早期生活史的补充机制、群落结构组成和生物多样性、渔业种群评估和管理、优势种演替等方面。然而, 对于该海域游泳动物群落的长周期变化和演变趋势的研究相对较少。为了研究长江口渔场游泳动物群落的年代际演变特征, 考虑到该海域游泳动物的繁殖、育幼、索饵等活动主要发生在春、夏、秋等3个季节, 为减少游泳动物补充量的变化对群落表征的影响, 同时需去除夏秋季东海北部大型水母暴发对游泳动物群落的影响[11, 12], 因此本研究选用群落季相较为稳定的1991— 2011年冬季的调查资料来分析各年代长江口渔场游泳动物种类组成、生物量、优势种、多样性和长度谱的变化特征, 研究造成其年代间优势种更替和群落差异性的原因, 从生态系统保护和渔业可持续利用角度提出管理建议。
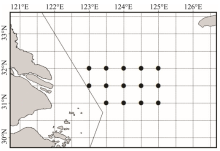
数据来源于1991年1月24— 26日、2001年12月25— 30日、2011年12月17— 20日在长江口渔场(31º 00′ ~32º 00′ N、125º 00′ E以西, 禁渔区线外的水域)进行的渔业资源调查; 调查站位设置如图1所示。1991年调查船为单拖调查船, 功率1 176 kW, 每站拖网1 h, 平均拖速3.5 kn, 平均袖网间距19.8 m, 囊网网目25 mm; 2001和2011年的调查船均采用220 kW双拖渔船, 每站拖网1 h, 拖网速度为3 kn, 网口宽度35 m, 网囊网目25 mm。每站所捕获样品随机取样1箱, 样品不满1箱时则全部取样, 然后带回实验室进行分类鉴定, 记录每种的重量和尾数。
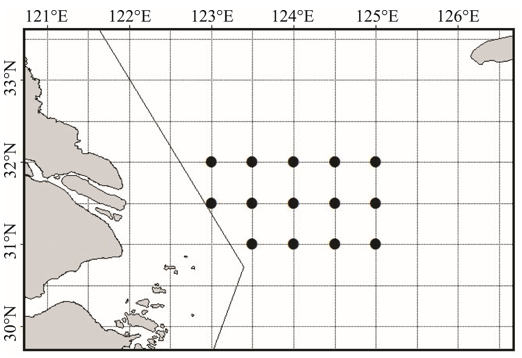 | 图1 长江口渔场调查站位示意图Fig. 1 Survey stations in Yangtze estuary fishing ground |
为了消除不同时段调查由于网口宽度、拖速与捕获率不同所产生的误差, 需要对两种网具进行标准化。考虑到不同网具的网目大小及其本身的材料等因素, 鱼类的逃逸率有差别, 因此单拖网的捕获率取0.5, 双拖网的捕获率取0.7; 对于不同调查网具的扫海面积, 由于拖网速度基本一致, 因此可以用网口宽度来代替扫海面积。网具标准化过程为:考虑到调查所使用网具的规格不同, 对游泳动物的捕获率[28]有差别, 因此以双拖网为基准, 对1991年单拖网的资源密度值进行标准化。数据标准化公式为:单拖网标准化数据=单拖网调查数据× (双拖网扫海面积/单拖网扫海面积)× (双拖网捕获率/单拖网捕获率)。
采用Margalef丰富度指数(D)、Shannon多样度指数(H′ )及Pielou均匀度指数(J′ )等[29, 30, 31]来研究不同年代该水域的游泳动物多样性。
用Bray-Curtis相异性指数来计算不同年份间游泳动物群落组成的差异, 其计算公式为:
式中:Xij、Xim分别为第i种游泳动物在j和m年调查中的渔获量, S为种类数。
所有数据输入Access数据库进行查询统计, 利用Prime 5.0软件进行多样性指数的计算, 并利用统计软件SPSS 18.0进行数据检验分析。优势种的确定采用Hill’ s N2指数[32], 可在Prime 5.0软件上进行计算。
根据每种游泳动物的总质量和总尾数, 计算平均体质量(W, g), 将平均个体质量转换成平均长度(L, cm), 公式为:L=(100× W)1/3, 长度谱变化组距为5 cm, 然后将该种类的网获尾数分配到相应的长度组中。以个体长度组平均每网每小时渔获总尾数(N)的自然对数为纵坐标, 长度组作为横坐标作两者的关系图, 分析3次调查中渔获物的长度谱变化。
不同生态类游泳动物单位捕捞努力量渔获量(CPUE)的年代际变化统计结果(表1)显示, 1991— 2011年, 长江口渔场游泳动物的重量CPUE表现出先急剧下降, 再中尺度回升的趋势; 尾数CPUE表现出先略有下降, 然后快速升高的趋势。造成重量CPUE和尾数CPUE变化趋势不一致的原因, 主要和游泳动物平均体重的变化有关, 可以看出:随着年代的推移, 平均体重先呈现急剧下降然后小幅下降的趋势。种类数方面, 各生态类的种类数均表现出升高的趋势, 但鱼类的增加幅度较小, 甲壳类和头足类的增加幅度较大。
| 表1 不同生态类游泳动物CPUE的年代际变化 Table 1 Inter-decadal changes of CPUE in different ecological categories of nekton |
根据对冬季长江口渔场游泳动物Hill’ s N2指数的计算结果, 1991年优势种为7种, 2001年优势种为9种, 2011年优势种为7种, 具体优势种见表2。由表2可见:1991年, 优势种依次为细点圆趾蟹、鳓、赤魟、细纹狮子鱼、三疣梭子蟹、龙头鱼、灰鲳; 2001年与1991年相比, 只有细点圆趾蟹和龙头鱼仍处在优势种之列, 新上升为优势种的7个种类为黄鲫、带鱼、银鲳、棘头梅童鱼、凤鲚、口虾蛄、虻鲉; 2011年与2001年相比, 龙头鱼、细点圆趾蟹、黄鲫、带鱼、凤鲚等5个种类仍处于优势种之列, 鳀和鹰爪虾新上升为优势种。综上所述, 后2个年份和1991年相比, 优势种发生了较大的更替, 共同优势种仅有2种。
| 表2 各年代游泳动物优势种组成 Table 2 Composition of dominant nekton species in different years |
由各年间游泳动物平均站位多样性指数的变化(表3)可见:随着年代的推移, 冬季长江口渔场游泳动物的站位平均种类数呈现持续升高趋势, 而站位平均丰富度指数、多样度指数和均匀度指数则呈现出先升高再降低的趋势。单因素方差分析(ANOVA)结果表明:各年份两两之间在种类数指标方面均没有显著性差异(P> 0.05), 但在丰富度指数、多样度指数和均匀度指数等3个指标方面均存在显著性差异(P< 0.05)。
| 表3 各年代游泳动物平均站位多样性指数的变化 Table 3 Inter-decadal changes of average nekton diversity index |
年间游泳动物群落的相异性分析表明, 1991和2001年、1991和2011年之间的平均相异性较高, 均超过了60%, 而2001和2011年之间的平均相异性较低, 为36.05%(表4)。对引起不同年间群落差异的百分比贡献率超过2%的主要种类进行统计, 结果(表4)表明:引起1991和2001年群落差异的主要种类有8种, 其中鱼类7种, 甲壳类1种; 引起1991和2011年群落差异的主要种类有10种, 其中鱼类8种, 甲壳类1种, 头足类1种; 引起2001和2011年群落差异的主要种类有12种, 其中鱼类6种, 甲壳类3种, 头足类3种。
| 表4 不同年代影响群落差异的主要种类百分比贡献率 Table 4 Contribution percentage of major species to community dissimilarity in different years (%) |
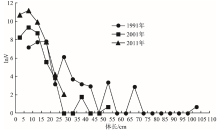
由冬季长江口渔场游泳动物长度谱的年代际变化(图2)可见:随着时间的推移, 游泳动物的长度谱呈现逐渐变窄的趋势, 长度谱的最大值先从1991年的102.5 cm减小到2001年的52.5 cm, 接着至2011年又减小到27.5 cm。同时, 在资源尾数方面, 长度20 cm以下的小个体随着时间的推进呈现出持续增加的趋势。这些都说明群落中的大个体高龄生物逐渐减少或消失, 而小个体低龄生物在群落中逐渐占据优势。趋势回归分析表明:各年代的趋势线呈线性回归, 渔获尾数的自然对数和体长呈负相关关系; 随着年代的推移, 趋势线的斜率绝对值和截距均呈增加的趋势。各年代长度谱的回归方程和相关参数见表5。
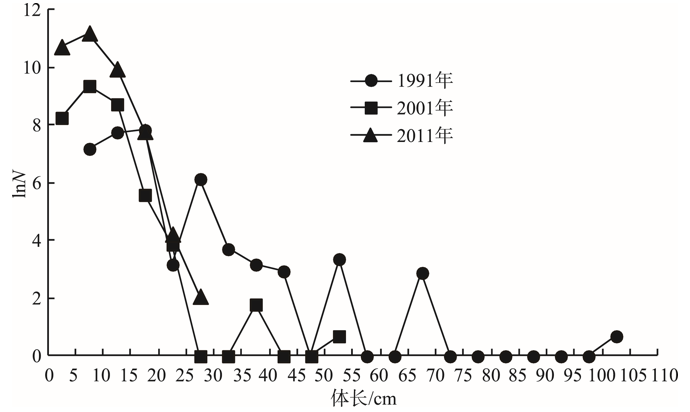 | 图2 冬季长江口渔场游泳动物长度谱的年代际变化Fig. 2 Inter-decadal changes of nekton size spectrum in Yangtze estuary fishing grounds in winter |
| 表5 各年代长度谱回归方程和相关参数 Table 5 Regression equation and correlation parameters of size spectra in different years |
从冬季长江口渔场游泳动物群落的演变来看, 1991— 2001年, 站位平均种类数、丰富度指数、多样度指数和均匀度指数均明显增加, 重量CPUE大幅降低, 尾数CPUE则略有降低; 2001— 2011年, 站位平均丰富度指数、多样度指数和均匀度指数均小幅减小, 但站位平均种类数略有增加, 重量CPUE和尾数CPUE大幅升高。21世纪初期与20世纪90年代初期相比, 多样性升高的可能原因如下:持续高强度的捕捞导致近海底层生物资源衰退, 大个体游泳动物减少, 饵料生物更加丰富, 因此小型游泳动物的捕食空间和饵料来源变大; 捕食压力的降低使得小型鱼类数量急剧增加, 成为控制群落能量流动的优势种, 进而改变了群落结构[20], 而中等程度的干扰水平能维持高的群落多样性。虽然有研究表明, 1982— 2005年东海北部鱼类群落总体多样性呈现降低的趋势[5], 但由于捕捞活动的不均匀性, 长江口近海渔场渔业资源的衰退可能导致重点捕捞海域向外海转移, 降低了该海域的捕捞压力; 同时, 高营养级底层鱼类多样性的降低可能使更低营养级的甲壳类和小型头足类多样性大幅升高[11]。因此, 21世纪初期长江口渔场游泳动物的多样性相对于20世纪90年代来说, 总体表现出升高的趋势。在尾数CPUE降低幅度较小的情况下, 游泳动物平均体重的快速下降是2001年的重量CPUE比1991年大幅降低的原因; 在游泳动物平均体重降低幅度较小的情况下, 尾数CPUE大幅回升是2011年重量CPUE较2001年有一定程度回升的原因。长江口渔场游泳动物群落的演变是东黄海游泳动物群落演变的缩影, 主要受东海区海洋渔业捕捞生产的影响, 捕捞压力会通过生物个体的变化传导至游泳动物群落使其发生结构演变。根据中国渔业统计结果, 1991、2001、2011年东海区的海洋捕捞产量分别为254.95× 104、612.71× 104、553.63× 104t, 而单位捕捞努力量渔获量分别为0.76、0.99、0.85 t/kW。由此也可以看出, 游泳动物群落的演变趋势主要受上述海洋捕捞生产相关因素的影响。
从优势种的变化来看, 1991年和后2个年份的优势种差异性较大, 2001和2011年的优势种重合度较高; 3个年份的共同优势种是细点圆趾蟹和龙头鱼。另外, 后2个年份还有3个种类共为优势种, 分别是黄鲫、带鱼、凤鲚。20世纪90年代以来, 随着长江口水域的大黄鱼、小黄鱼(Larimichthys polyactis)、鳓等主要经济鱼类的持续衰退, 龙头鱼、黄鲫和凤鲚等次优经济鱼类已成为当前该海域游泳动物群落的主要优势种和冬春季的重要渔业捕捞对象[4, 6]。随着年代的推移, 长江口渔场游泳动物的群落相异性和引起群落差异的主要种类数均呈增加的趋势, 并且生态类群也逐渐由鱼类和甲壳类扩展到鱼类、甲壳类、头足类, 甲壳类和头足类在群落中的重要程度逐渐增加。这表明长江口渔场的游泳动物群落结构的年代际变化较为明显, 且前期的变化程度相对较剧烈, 后期的变化程度较舒缓, 其原因一方面可能与东海区1991— 2001年的捕捞努力量增加较快(1991和2001年分别为334.75× 104、615.94× 104kW)、2001— 2011年增加幅度较小(2011年为649.80× 104kW)有关; 另一方面, 三峡工程的成功截流蓄水, 也改变了长江径流原有的季节分配格局, 使冬季枯水期径流量下降幅度减小, 而长江径流量的大小对于河口的水文物理条件有显著的影响, 制约着生物群体的变迁, 影响着游泳动物在产卵场、索饵场和越冬场之间的洄游行动[6, 18]。
游泳动物群落结构的变化可以通过长度谱的变化来衡量。长度谱的减小, 使其所占的生态位降低, 对群落能流的贡献减小, 不存在绝对的优势种。为了维持群落的平衡, 群落内部自我调节, 种类更替加速, 允许更多的种类能够生活在该生态系统内, 群落内优势种的优势度达到相对均衡, 因此物种的替换所达到的适应性更为有效[33]。1991— 2011年, 长江口游泳动物长度谱呈现小个体逐渐增加、大个体持续减少的趋势。这种长度谱的变化趋势主要是由于群落中生物种类的演替和群体年龄结构的降低引起的。如1991年的鳓、赤魟、细纹狮子鱼、灰鲳等优势种至2001年后已不在群落中占优势, 代之以棘头梅童鱼、凤鲚、口虾蛄、虻鲉、鳀、鹰爪虾等小型鱼类或甲壳类为优势种。同时, 一些主要种类如带鱼、小黄鱼、日本鲭(Scomber japonicus)等也表现出持续的低龄化现象, 群体组成以当龄鱼和1龄鱼为主, 高龄鱼基本消失[34, 35]。由此说明, 自20世纪90年代以来, 长江口渔场游泳动物群落发生了较大的演替现象, 游泳动物种类正由经济价值高、个体大、年龄结构复杂和营养层次高的种类向经济价值低、个体小、年龄结构简单和营养层次低的类群演变, 并且在此过程中种类的更替速度呈加快的趋势。目前, 该水域的游泳动物群落维持在低营养层次的动态平衡状态。出现这种现象的原因一方面与小型鱼类的数量增加有关, 另一方面也与大型底层和中上层经济鱼类在高强度的捕捞压力下的年龄结构变简单、性成熟年龄提前、从K选择型向r选择型转变[34, 35, 36]等自身调节机制有关。在国内其他河口和渔业资源研究中也有类似结果出现[37, 38]。从生态保护和渔业可持续利用的角度出发, 为了使该海域的游泳动物群落恢复至正常水平, 渔业行政管理部门除了继续实施伏季休渔制度, 严格执行渔业法规定等相关管理措施外, 还要合理控制或降低捕捞努力量, 减小对经济种类的捕捞强度、适度增加对甲壳类和小型头足类的利用程度, 特别是针对个别经济种类在其主要产卵场或育幼场设立特别保护区或特定休渔期, 对条件成熟的鱼种实行TAC(Total Allowable Catch)管理。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|