

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1980—2014年中亚地区植被净初级生产力对气候和CO2变化的响应
[朱士华1, 2, 3  , 艳燕
, 艳燕2, 3 , 邵华2, * , 李超凡2, 4 ]
, 艳燕, 李超凡|
|


第一作者简介:朱士华(1989- ),男,博士,研究方向为生态模型、全球变化生态学。E-mail:zshcare@ foxmail.com
*通信作者简介:邵华(1969- ),女,副研究员,研究方向为生态学。E-mail:shaohua@ms.xjb.ac.cn
中亚干旱区分布着世界80%以上的温带荒漠,受气候变化影响显著。论文首先收集实验观测数据验证了干旱区生态系统模型(AEM),然后运用AEM开展数值模拟实验量化研究了1980—2014年中亚净初级生产力(NPP)的时空格局,评估了不同环境因子(降水、温度、CO2)的相对贡献率及其交互效应。结果表明:过去35 a中亚干旱区年均NPP总量为1 125±129 Tg C(1 T=1012)或218±25 g C/m2。哈萨克斯坦北部地区年NPP值较高(349±39 g C/m2),而南疆地区年NPP值较低(123±45 g C/m2)。1980—2014年间,中亚NPP总体呈减少趋势 [-0.71 g C/(m2·a)],南疆极端干旱区的NPP降低最为显著 [-2.05 g C/(m2·a)]。相较于1980—1984年NPP均值,在1985—2014年中亚区域NPP总体降低了118 Tg(-10%)。其中CO2施肥效应促进NPP增加了99.7 Tg (+8%),气温升高的正效应促进NPP增加了35.4 Tg(+2%),而降水减少导致NPP降低了221 Tg(-18%)。研究区内9%的地区的NPP主要控制因子为温度,主要分布在天山和哈萨克斯坦北部等高纬高寒地区。降水主控区面积占整个研究区的69%,主要分布在荒漠平原特别是南疆等植被受水分限制的区域。CO2主控区占研究区面积的20%,主要分布在天山中山带森林区和低海拔地区等水热条件好的区域。研究表明新疆南部地区是中亚的关键生态脆弱区,其生态安全面临着气候变化的挑战,但21世纪的升温不大可能因刺激自养呼吸而对中亚区域NPP造成显著影响。
More than 80% of world’s temperate desert locates in Central Asia, dryland that experienced strong warming and significant changes in precipitation pattern in recent decades. The objectives of this study was to quantify spatio-temporal patterns of net primary productivity in Central Asia over the past 35 years, and investigate the relative contribution and interactive effect of climate controls including temperature, precipitation and CO2, using the Arid Ecosystem Model (AEM), which performed well in predicting the dryland ecosystems’ responses to climate changes according to our evaluation. The results show the 35-year averaged annual NPP of Central Asia amounted to 1 125±129 Tg C/a (1 T=1012), or 218±24 g C /(m2·a), with an overall declining trend of 0.71 g C/(m2·a). The northern Kazakhstan had relatively high NPP of 349±39 g C/(m2·a), while the southern Xinjiang of China had relatively low NPP of 123±45 g C/(m2·a). During the last 35 years, the NPP of the southern Xinjiang subregion declined significantly with a trend of -2.05 g C/(m2·a). Comparing the NPP during 1985-2014 to the NPP during 1980-1984, we found the regional NPP decreased 118 Tg or 10%, with positive contribution of 35.4 Tg or +2% from temperature change, positive contribution of 99.7 Tg or +8% from CO2 change, negative contribution of 221 Tg or -18% from precipitation change. The temperature was the dominant factor on NPP in 9% of the study area, mainly in the Tianshan Mountains and northern Kazakhstan, where the temperature increased by 0.03 ℃/a during 1980-2014. Precipitation was the dominant factor on NPP in 69% of the study area, mainly in the desert subregion and the dryland of southern Xinjiang of China, where the vegetation was limited by water. CO2 was the dominant factor on NPP in 20% of the study area, mainly in the lower altitude regions of Tianshan Mountains, where the hydrothermal condition was suitable for vegetation growth. This study shows that the southern Xinjiang of China is the fragile eco-environmental subregion in Central Asia, whose ecosystem security is facing the challenges of climate change. However, increasing temperature is unlikely to stimulate autotrophic respiration and cause a significant impact on regional NPP in Central Asia.
中亚地处“ 新丝绸之路” 核心地带, 包括哈萨克斯坦、乌兹别克斯坦、土库曼斯坦、吉尔吉斯斯坦、塔吉克斯坦以及中国新疆地区, 总面积500多万km2, 拥有世界80%以上的温带荒漠, 正面临着气候变化的严重威胁[1]。由于地理异质性显著, 气候系统以及土地利用过程复杂[2], 生态系统对全球变化的响应非常敏感[3]。近30 a中亚升温速度达0.4 ℃/10 a, 显著高于北半球陆地(0.3 ℃/10 a)和周边地区的升温速率, 并在空间上呈现中部升温快和升温速率从西北到东南递减的格局[4]。同一时期, 中亚西北地区发生了严重的干旱, 而中亚东部新疆地区降水显著增加[5], 中亚降水表现出强烈的空间异质性[1]。
气候变化及其对中亚独特的温带荒漠生态系统造成的环境压力, 威胁着“ 新丝绸之路” 核心地区的生态安全。
植被净初级生产力(Net Primary Production, NPP)指绿色植物在单位时间和单位面积上积累的有机碳总量, 它不仅是生态系统中物质和能量运转研究的基础, 还是判定生态系统碳收支和调节生态系统过程的主要因子[6]。目前, 中亚植被NPP的研究匮乏, 除全球尺度的NPP研究外鲜有涉及, 李超凡等[7]基于遥感数据对近20 a中亚5国净初级生产力的格局做了阐述, 但遥感数据忽略了时空尺度的变化机理。张建财等[8]利用LPJ模型估算了亚洲中部干旱区的植被NPP以及蒸散时空格局, 初步统计了中亚干旱区生态系统的生产力变化情况, 但模型未涉及干旱区植被特殊生理生态特征及其对气候变化的响应。因此, 揭示植被NPP的时空格局并量化其主要气候控制因子, 对评估中亚生态安全和提出适应性对策具有重要意义。AEM(Arid Ecosystem Model)[9]是针对中亚干旱区特殊的生理生态过程(高根冠比、垂直根分布水分吸收模式等)发展的生态过程模型, 可模拟温带荒漠植被的结构和功能以及生态系统碳水循环的复杂相互作用[10]。AEM被应用于中亚碳库时空格局评估[11], 但是仍需要评估AEM预测中亚植被对CO2、温度、降水等环境因子的响应能力, 以进一步量化分析不同气候因子对植被NPP变化的相对贡献率。因此, 本研究首先收集实验观测数据验证同等试验条件下AEM对植被-气候响应动态的预测能力; 继而运用AEM开展数值模拟实验, 量化研究1980— 2014年中亚植被NPP的时空格局, 评估不同环境因子(降水、温度、CO2)的相对贡献率及其交互效应, 以NPP变化为指标识别气候变化背景下的中亚生态脆弱区。
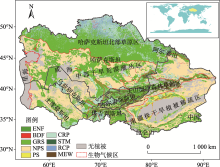
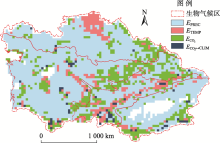
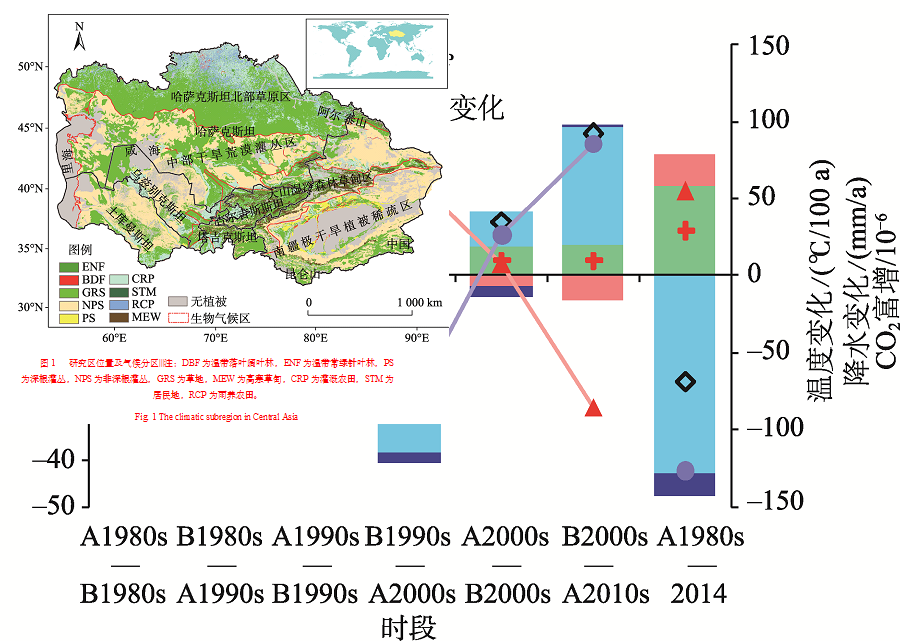
中亚干旱区(34.3° ~55.4° N, 46.5° ~96.4° E)处于亚欧大陆腹地, 其地势东南高、西北低, 具有典型的山盆结构特征, 该区域独特的山地-绿洲-荒漠生态系统格局使其对全球变化响应特殊而复杂(图1)。为进一步分析不同气候带控制下的中亚地区植被净初级生产力格局, 依据Hu等[4]的标准将中亚地区分为4个地理气候区, 分别为哈萨克斯坦北部半干旱草原区、中部干旱荒漠灌丛区、天山湿冷森林-草甸区(常绿针叶林与高山草甸为主)、中国南疆极干旱植被稀疏区。
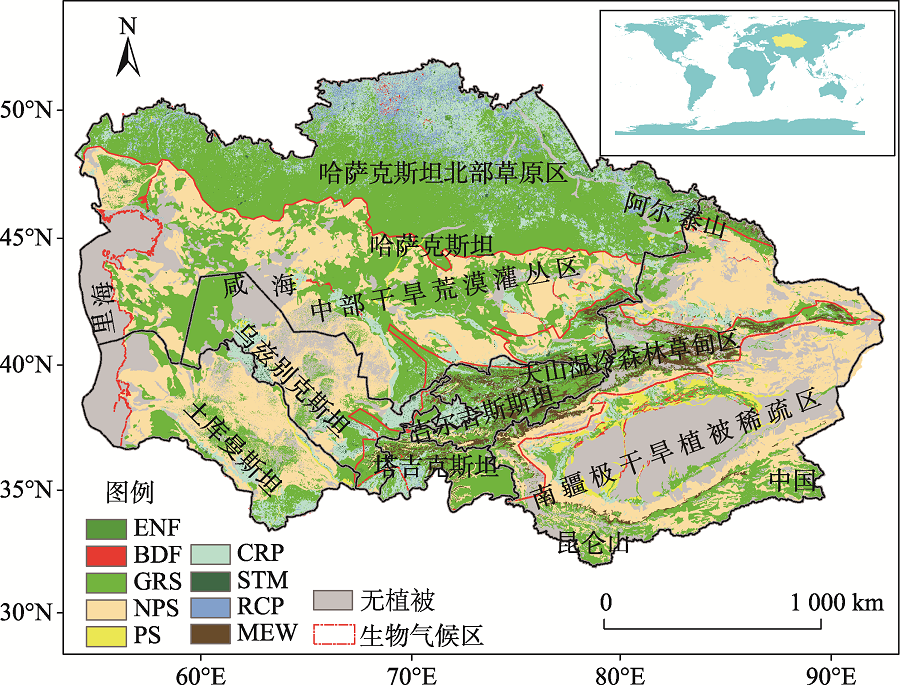 | 图1 研究区位置及气候分区注:DBF为温带落叶阔叶林, ENF为温带常绿针叶林, PS为深根灌丛, NPS为非深根灌丛, GRS为草地, MEW为高寒草甸, CRP为灌溉农田, STM为居民地, RCP为雨养农田。Fig. 1 The climatic subregion in Central Asia |
中亚地区主要受北半球西风环流控制并依赖于大西洋湿润气团带来降水, 其湿度年际变化格局同亚洲季风区明显不同[12]。来自北方的极地急流和来自南方的阿拉伯海气流扰动降水季节格局, 增加了气候的不确定性。再加上绿洲湿岛效应以及全球变暖背景下副热带急流北移效应共同造成了中亚干旱区复杂的气候变化格局。
1.2.1 模型验证和比较
AEM模型可动态模拟日、年尺度的站点和区域生态系统碳、氮、水通量和储量, 相较于其他生态过程模型, AEM模型针对干旱区植被特殊生理生态过程, 细化了4个关键机制的模拟:群落和个体形态结构对生态过程影响的模拟; 地下水对深根植物生长影响过程的模拟; 地下水-土壤-根系-冠层连续体之间水循环关键过程的模拟; 耐旱植被具有高效节水光合能力的模拟[9, 10]。AEM的基本模拟单位为植被功能类型(plant functional type, PFT), 对中亚干旱区9种典型植被功能类型进行了本地参数化, 分别是温带落叶阔叶林、温带常绿针叶林、深根灌丛、非深根灌丛、草地、高寒草甸、灌溉农田、居民地和雨养农田[10]。然而尚需检验在这些模型参数驱动下, AEM能否准确预测不同PFT对各种气候因子的响应动态。
为此, 本研究检索了23个环境因子控制实验研究(表1), 针对这些站点的实验设计, 研究设计同样的气候环境(初始场相同, 包括温度、降水、相对湿度等)和试验条件(不同环境梯度), 并定量开展AEM模拟, 将模型预测的NPP响应(如CO2倍增情景下NPP的增长百分比)同实验观测值进行比较, 以校验AEM模型以及不同植被功能型的气候敏感性。此外, 在区域尺度上, 将AEM模拟的中亚植被多年平均NPP同遥感反演的NPP进行了点对点的对比。其中, 1980— 1999年的遥感NPP数据来自基于AVHRR影像的GLO-PEM(Global Production Efficiency Model)模型反演产品(ftp://ftp.glcf.umd.edu/glcf/GLOPEM/), 2000年及以后的遥感NPP数据基于MODIS (Moderate Resolution Ima-ging Spectroradiometer) 遥感数据产品 (https://ladsweb.nascom.nasa.gov/data/search.html)。
| 表1 所收集的气候变化(CO2、温度、降水)对植被生产力影响的实验研究列表 Table 1 A list of experiments on the effects of climate factors on NPP |
1.2.2 中亚区域模拟的数据源
1980— 2014年逐日尺度40 km × 40 km分辨率气象数据来自美国国家环境预报中心的CFSR气候再分析数据(包括日降水、最高温度、最低温度、平均温度、太阳短波辐射、相对湿度, http://rda.ucar.edu/pub/cfsr.html)。地理基础数据主要包括地形数据(高程、坡度、坡向)、植被类型数据、土壤数据(粘土、粉砂和砂含量、容重、pH值等)、地下水位等。所有相对高分辨率(< 40 km)的源数据均采用栅格空间计算, 聚合成40 km × 40 km的统一分辨率开展模拟。植被NPP为总初级生产力(GPP)和自养呼吸(RA)的差值。为研究NPP变化机理, 本模拟研究输出了研究区每个40 km × 40 km格点内每种PFT类型从1980到2014年每日的NPP、GPP和RA数据。
1.2.3 气候因子效应分析的数值情景实验设计和分析方法
为量化不同气候因子对1980— 2014年NPP变化的相对贡献率, 本研究采用AEM模型开展数值模拟实验。共设计了5种实验情景以开展因子分析(表2)。SOVERALL模拟真实情景, 即气候变化以及多种气候因子交互作用下的NPP动态;
| 表2 模拟实验情景设计 Table 2 Scenario design |
即:
EPREC = NPPL, PREC - NPPE, PREC(2)
ETEMP = NPPL, TEMP - NPPE, TEMP(3)
式中:
站点尺度验证表明AEM可准确预测气候因子梯度变化对中亚典型PFT类型NPP的影响。其中, 二氧化碳富增实验中, R2=0.86, RMSE=0.07; 温度站点验证中R2=0.52, RMSE=0.19; 降水站点验证实验中R2=0.98, RMSE=0.04(图2)。所有气候因子验证均显示AEM模拟与观测的NPP显著相关(P < 0.01)。AEM可正确预测不同植被对气候因子敏感性的差异:不同PFT对CO2富增的敏感性表现为森林< SRB< GRS, 且森林中ENF> DBF; GRS对温度的敏感性要小于SRB和ENF; DBF对降水的敏感性要小于灌木。这些差别与其不同PFT的生存环境相关。例如, 灌木一般生长在干旱少雨的荒漠, 受降水胁迫影响, 其对降水敏感性要大于生存在降水相对较多区域的阔叶林。
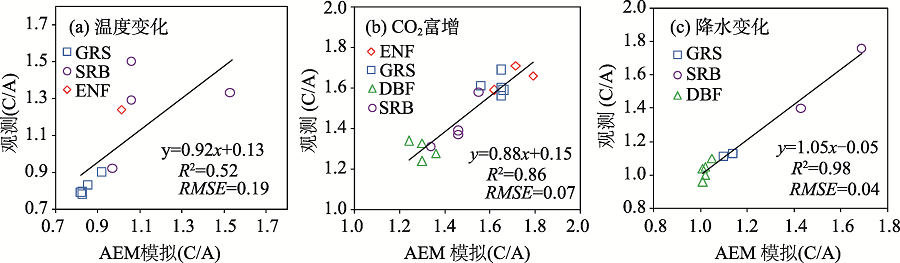 | 图2 中亚PFT对气候变化的敏感性注:C/A即气候因子变化后植被NPP同当前NPP(对照)的比值; 所有气候因子验证均显示AEM模拟与观测的NPP显著相关(P < 0.01)。Fig. 2 The effects of climate change on the NPP of major PFTs in Central Asia |
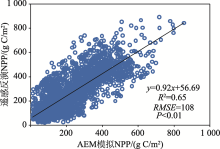
区域尺度评估表明AEM预测的NPP与遥感影像获取的NPP格局具有高度的一致性(图3), 二者线性拟合曲线的斜率为0.92, R2=0.65, RMSE为108 g C/m2(P < 0.01)。该结果进一步表明AEM对中亚NPP的模拟结果同遥感反演产品总体上可以相互印证。
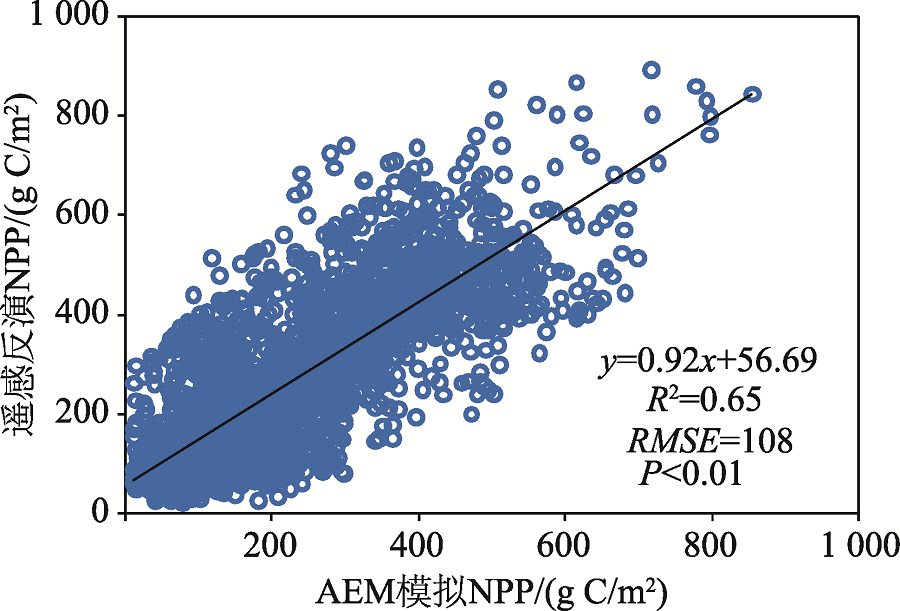 | 图3 AEM模拟的多年平均NPP值与遥感反演NPP间的点对点比较(1980— 2010年)Fig. 3 The comparison between the long-term mean NPP based on AEM simulation and remote sensing models (MODIS and GLO-PEM) during 1980-2010 |
研究结果表明:SOVERALL情景下, 中亚干旱区植被平均NPP总量为1 125± 129 Tg(或218± 25 g C/m2), GPP总量为1 937 Tg(或375g ± 39 C/m2), RA总量为1 002 Tg(或194± 39 g C/m2)。近35 a中亚NPP以0.71 g C/(m2· a)的总体趋势减少, 年际波动显著(图4)。1980— 1997年间NPP趋势平稳, 1998— 2008年间NPP起伏较大, 2001、2006、2008年有3次较为明显的NPP低谷, 与拉尼娜现象发生的时间吻合;
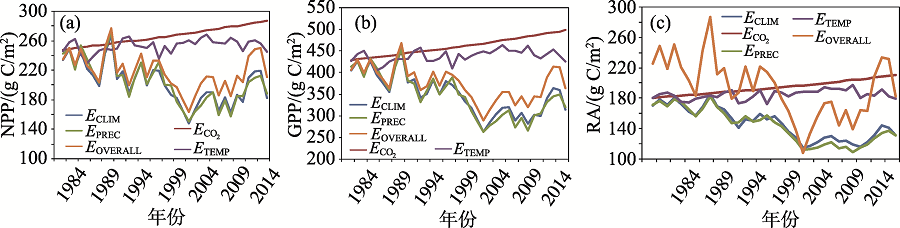 | 图4 不同情景下中亚地区植被NPP、GPP和RA年际动态(1980— 2014年)Fig. 4 The temporal pattern of NPP, GPP and RA in Central Asia from 1980 to 2014 |
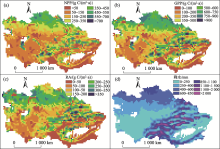
植被生产力(NPP和GPP)的空间分布同降水的空间格局密切相关(图5), 其高值区主要分布在水源丰富的哈萨克斯坦北部草原 [平均NPP=349± 39 g C/(m2· a), GPP = 577± 61 g C/(m2· a)] 和天山西北迎风坡中山带 [天山分区的平均NPP=236± 22 g C/(m2· a), GPP = 378± 32 g C/(m2· a)]。低生产力地区主要包括中亚西南荒漠 [荒漠分区的平均NPP=175± 23 g C/(m2· a), GPP=321± 34 g C/(m2· a)] 以及南疆地区 [平均NPP=123± 45 g C/(m2· a), GPP=217± 75 g C/(m2· a)], 这些地区属于典型的温带荒漠气候, 降水稀少、蒸发强烈、水分胁迫严重, 绿洲灌溉农田对其NPP有很大贡献(图5中绿色破碎斑块)。RA的空间格局也总体受降水影响, 但是在哈萨克斯坦北部草原靠近边界一侧降水较高(> 600 mm/a)的区域RA却较低, 可能是处于高纬高寒地区(50° N, 平均温度 < 2 ℃)、受到低温抑制的缘故。
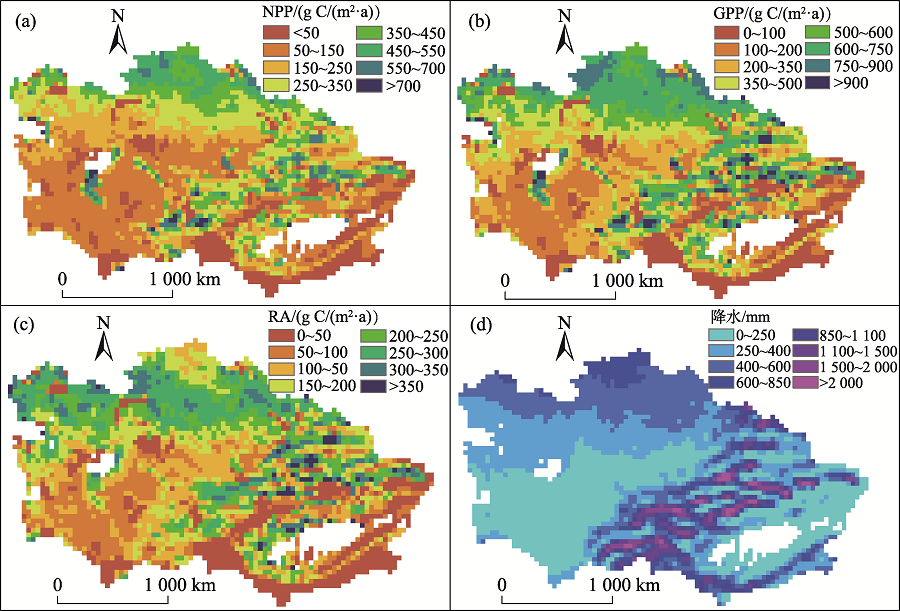 | 图5 中亚干旱区NPP、GPP、RA以及降水的空间格局(1980— 2014年)Fig. 5 The spatial pattern of NPP, GPP, RA and precipitation in Central Asia from 1980 to 2014 |
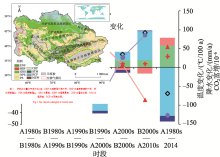
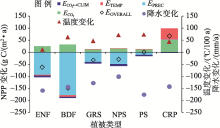
本文分析表明不同时段内环境因子的变化对NPP的影响不同(图6)。全球大气CO2浓度持续升高, CO2施肥效应促进中亚NPP以约6 g C/(m2· a)的速率稳定增长, 35 a增加了100 Tg C。然而降水的下降导致NPP在整个20世纪80年代到21世纪初的各时间段上都呈下降趋势。虽然在2000年代末后降水有所上升, 但过去35 a的降水变化总体上导致中亚的年NPP减少221 Tg C。相对于降水和CO2同NPP之间明显的正相关效应, 温度变化对NPP的影响更复杂。
 | 图6 不同环境因子对NPP的个体以及交互效应注:EOVERALL反映气候变化对NPP的总体影响; A1980s指1980— 1984年, B1980s指1985— 1989年, 其余类推。Fig. 6 The individual effect and interactive effects of different environment factors on NPP |
在1995— 1999至2000— 2004年以及2005— 2009至2010— 2014年期间, 温度变化同NPP动态为正相关, 其他时期则表现为负相关。整个研究段内中亚气温的上升促进了区域年NPP增长约35 Tg C。因子分析结果显示, CO2上升同气候变化(包括温度和降水等)间的交互作用减少了NPP, 特别是在21世纪初。这段时期中亚处于近百年的一个相对干旱期。在各种因子的综合作用下, 中亚NPP以21世纪初为界, 先持续减少, 然后反弹, 总体上下降了118 Tg C。这同降水影响下的NPP格局基本一致, 可见降水是中亚植被生产力的主控因子。
空间上, 不同区域的植被NPP主控因子不同(图7)。约69%的研究区主要受降水控制, 这些区域覆盖了中亚荒漠生态系统, 以及近35 a发生严重干旱的西北哈萨克斯坦草原区[4]。CO2施肥效应的主控区占研究区总面积的20%, 主要分布在天山中山带森林区和低海拔绿洲农田等水热条件较好的区域。温度主控区占研究区总面积的9%, 主要分布在哈萨克斯坦北部草原高纬度地区和天山、昆仑山高海拔地区, 都属于高寒地区。在以上3种气候因子影响力交界的昆仑山区零散分布有少量地区(占中亚2%)受CO2和气候因子交互作用影响。
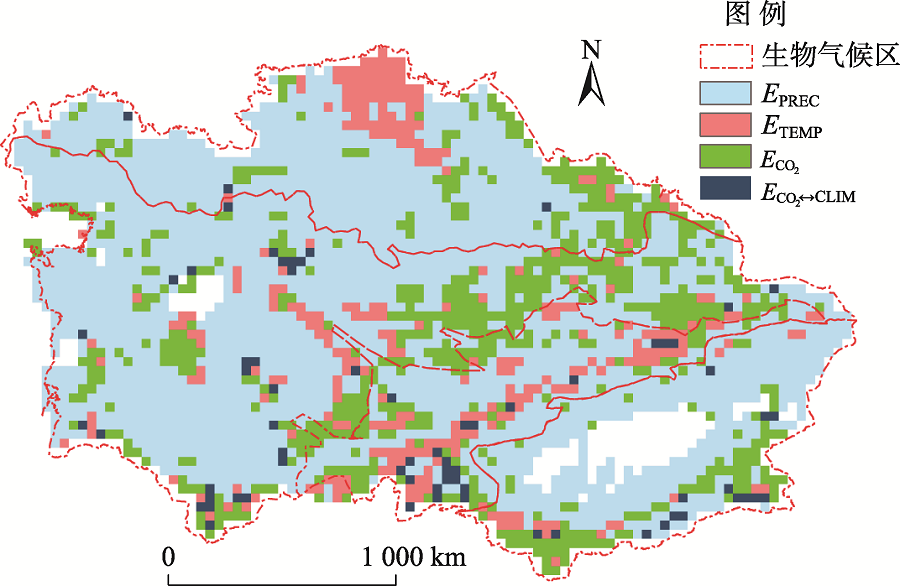 | 图7 中亚干旱区主控因子空间格局Fig. 7 The spatial pattern of major controls over the NPP in Central Asia |
通过分析ENF、BDF、GRS、NPS、PS和CRP六种主要的中亚PFT对环境因子的响应结果(图8)发现水肥条件良好的灌溉农田(CRP)对降水变化不敏感, 其NPP受到温度升高和CO2富增的促进。其他PFT类型的NPP均受中亚降水减少的影响而下降。其中阔叶林、针叶林因自身较高的NPP导致NPP下降的绝对值也最大。对于深根灌木和非深根灌木, 由于深根灌木的根系可以有效利用地下水, 所以对降水效应的敏感性要小于非深根灌木。除了CRP以外, 气温对各自然植被类型的影响均较小。CO2和气候因子交互作用对所有的植被类型均为负效应。
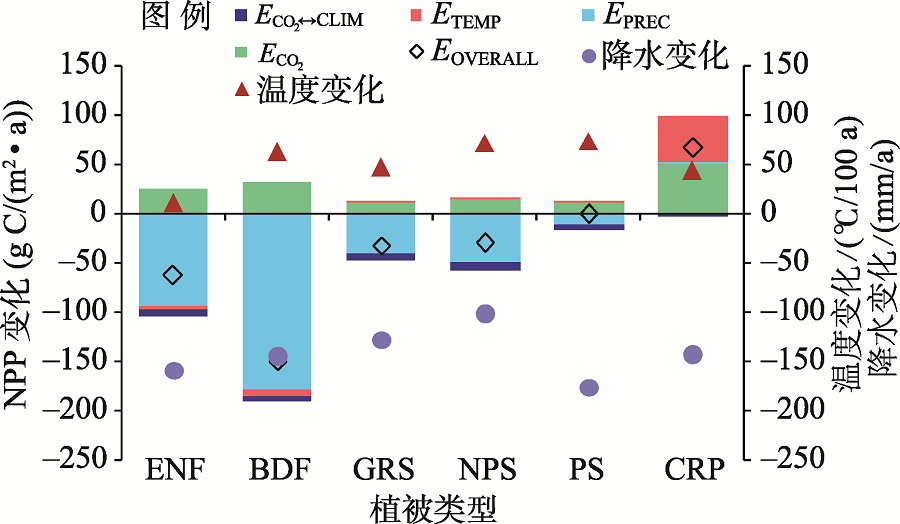 | 图8 不同植被类型对气候变化的响应Fig. 8 The responses of various PFTs to different climate factors |
研究发现中亚地区植被总NPP(1.13± 0.13 Pg C/a, 1 P = 1015)同澳洲植被NPP(1.86± 0.90 Pg C/a[36]或0.8~1.1 Pg C/a[37])相当, 约等于中国植被NPP(1.8~4.7 Pg C/a, 平均3.18± 0.84 Pg C/a)的1/3[38, 39, 40, 41, 42]、全球植被NPP(39.9~80.5 Pg C/a)的1.4%~2.8%[43, 44, 45, 46]。中亚地区的植被NPP总量对全球生态系统而言不可忽视。
此外, 因其所处的欧亚内陆中高纬地区是全球变暖的热点区域, 植被NPP的变化动态重要且独特[4]。温带地区的暖化一般有利于植物的光合作用和生产力提高。研究表明中国植被NPP在1982— 1999年间增加了24.2%, 近一半归因于气候变化[40]。印度1981— 2000年间NPP以8.5%/10 a的速度增加[47]。俄罗斯在1975— 2004年间88%的区域出现植被生产力增长[48], 且俄布里亚特大部分地区NPP与温度呈现显著正相关性[49]。但与周边地区不同, 我们发现中亚植被NPP在过去35 a下降了-10%(118 Tg C/a), 并主要损失于1980— 2000年间。分析显示中亚地区的植被NPP变化主要受降水影响(图7), 尤其是20世纪80年代中期到21世纪初的持续干旱(可能与La Niñ a现象相关)造成了中亚5国的荒漠和草原区(如图尔盖高原)NPP显著下降[11], 而气温上升也可能进一步加剧了干旱区(如南部新疆)的水分胁迫, 从而导致中亚植被退化[11]。
研究显示除北哈萨克斯坦高纬地区、高海拔山区以及降水条件较好的天山西坡和北坡外, 中亚大部地区植被NPP主要受降水控制 [图4(a)、图7]。这与Zhou等[50]观察到中亚大部分区域的植被NPP在20世纪90年代以来主要受降水影响的结论一致。Eisfelder 等[51]的研究也表明哈萨克斯坦的夏季植被NPP主要受降水控制, 但是有2个月的滞后效应。Gang等[52]的研究表明1981— 2010年间中国、北美、澳洲和欧洲的草原和荒漠植被NPP同降水变化显著正相关, 对气温的响应不显著或负相关。将SOVERALL同SPREC情景模拟结果对比, 可以发现不但其GPP的年际波动高度一致(R2=0.86, P< 0.01), 而且其RA的波动也基本相同(R2=0.70, P< 0.01)。虽然同大多数生态过程模型一样, AEM中的自养呼吸仅是温度和生物量的正相关函数:RA= f(TEMP) × Biomass[10], 但本文结果表明, SOVERALL情景下RA同降水的相关性(R2=0.73, P< 0.01)远高于同温度的相关性(R2=0.18, P> 0.01)。实验观测也显示荒漠生态系统的RA主要受水分影响[53], 而湿润区森林的RA(即使在热带)也对温度非常敏感[54]。这是因为干旱区植被对水分变化很敏感, 以至于水分主控下的生物量波动对RA的影响掩盖了气温对RA的直接效应。以下3个机制可解释相对于森林, 干旱区植被生物量对降水有较高的敏感性:1)中亚干旱区植被以一年生为主, 短命植物决定了温带荒漠群落约75%的生物量及其碳/水动态[55, 56], 因此中亚干旱生态系统生物量同当年生长期的气候条件(特别是降水)密切相关。与之对比, 湿润/半湿润地带性植被生物量是多年积累的结果, 受年内气候影响小, 因此其RA对温度更为敏感。本文模拟结果表明森林RA与气温的相关性(R2=0.34, P< 0.01)远高于荒漠草原(R2=0.16, P> 0.01)。2)多年生荒漠灌木可通过休眠应对干旱胁迫, 此时单位生物量的呼吸速率大为降低, 抵消了高温的影响[57]。3)升温虽然能刺激RA上升, 但也会增强水分胁迫而抑制生物量的增长。以上分析意味着, 中亚干旱区在21世纪的快速升温[4]可能不会因刺激RA而对其生态系统NPP产生显著的直接影响。在对中亚干旱区的生态模拟和气候效应评估中更应该关注因温度上升导致的潜在蒸散及水分胁迫增强对NPP造成的间接影响。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|